Úvod
Kreativní poznávání hraje důležitou roli v umění, v invenci a inovaci, stejně jako v každodenním životě (Runco a Richards, 1997). Přestože kreativita byla považována za jedinečnou lidskou kapacitu, spontánně kreativní chování (např. bylo prokázáno, že se vyskytují i u jiných lidí (Byrne a Bates, 2007), což dále potvrzuje hluboké evoluční a biologické kořeny kreativity. Kreativita byla definována jako schopnost produkovat odpovědi, které jsou jak román (tj., originální, vzácné a nečekané) a vhodná (tj. adaptivní a užitečné, v závislosti na úkolu omezení) (Sternberg a Lubart, 1999). Jeden z psychometrických přístupů k měření kreativity zahrnuje úkoly divergentního myšlení (DT), ve kterých jsou účastníci požádáni, aby reagovali na daný problém s více řešeními (Dietrich a Kanso, 2010). Testy DT obecně měří různé aspekty kreativity, včetně tvůrčí plynulosti ,flexibility a originality (Torrance, 1974). Ačkoli tedy nejsou synonymem kreativity, úkoly DT poskytují strukturovaná a objektivní měření kreativity (Sternberg a Lubart, 1999; Jung a kol., 2009). Zejména se ukázalo, že skóre úkolů DT pozitivně koreluje s ekologicky platnými měřítky tvůrčího úspěchu (Carson et al ., 2005) a také s vlastní kreativitou (Furnham a Bachtiar, 2008).
předchozí studie poukazovaly na zapojení dopaminergního (DA) systému do kreativity (Heilman et al., 2003; Flaherty, 2005; Takeuchi a kol., 2010). Takeuchi et al. (2010) zjistili, individuální rozdíly v tvořivosti, měřeno DT, být pozitivně koreluje s šedé hmoty v DA systém regiony, včetně dorzolaterální prefrontální kůra, bilaterální bazálních gangliích, substantia nigra a ventrální tegmentální oblasti. Navíc několik genetických studií ukázalo vztah mezi DT a dopaminovou neurotransmisí (Reuter et al ., 2006; De Manzano et al., 2010; Runco a kol., 2011). Například Reuter a kol. (2006) zjistil, že kreativita, měřená úkoly DT zahrnujícími figurální i verbální kreativitu, je významně spojena s polymorfismy genu receptoru dopaminu D2 (DRD2). Navíc několik studií zjistilo, že kortikální dopamin je zapojen do kognitivní flexibility(Frank, 2005; ochlazuje, 2008; Garcia-Garcia a kol., 2010), jedna z hlavních složek DT.
jedním zajímavým da kandidátským genem pro kreativitu je gen receptoru dopaminu D4 (DRD4). Receptor DRD4 je jedním z pěti dopaminových receptorů a hraje důležitou roli při zprostředkování synaptické dopaminové signalizace. Gen je charakterizován variabilním počtem tandemových opakování 48 párů bází (48 – BP VNTR) umístěných v kódující oblasti třetího exonu. DRD4 48 – BP VNTR polymorfismus se pohybuje od 2 do 11 opakování napříč jednotlivci (Asghari et al., 1995; Cravchik a Goldman, 2000), kde 4-opakování (4R) je nejčastější opakovat v Kavkazské populace a 7-opakování (7R) je druhá nejčastější varianta (Chang et al., 1996; Ding et al., 2002). Zajímavé je, že 7R byl dříve spojován s kreativním chováním v reálném životě, jako je osobnostní rys hledající novinku (Ebstein et al., 1996). Hledání novosti-tendence k průzkumné činnosti – je skutečně považována za jednu z charakteristik kreativních lidí (Chavez-Eakle et al., 2006; Drago a kol., 2009). Ačkoli důkazy o spojení mezi DRD4 7R a hledáním novinek jsou nekonzistentní, o čemž svědčí několik zpráv o metaanalýze (Kluger et al., 2002; Munafò et al., 2008), mnoho studií zjistilo významnou souvislost mezi hledáním novinek a alelou 7R DRD4 (Ebstein et al ., 1996; Benjamin a kol., 2000; Keltikangas-Järvinen et al., 2002; Becker et al., 2005). Roussos et al. (2009) nedávno navrhli, že rozdíly v měření hledání novinek pomocí měřítek self-report mohou odpovídat za nekonzistentní výsledky mezi polymorfismem 7R a hledáním novinek. V souladu s tím, to bylo opakovaně zjištěno, že DA systém hraje významnou roli v osobnostní rys novosti-seeking (Flaherty, 2005; Schweizer, 2006), dále potvrzující potenciální roli DRD4 v kreativitě.
Nicméně, v kontrastu k roli DRD4 48-bp VNTR 7R ve novinkou-převíjení, nedávné důkazy naznačují, že 7R je ve skutečnosti spojena s poruchou kognitivní flexibilitu, jedním z hlavních aspektů tvořivosti (Strobel et al., 2004; Congdon a kol., 2008). Flexibilita zahrnuje schopnost provést alternativní odpověď po úspěšné inhibici aktuální odpovědi. Dosavadní zjištění týkající se souvislosti mezi inhibicí a DRD4 jsou smíšená. Congdon a kol. (2008) zjistili, že účastníci s alelou 7R DRD4 vykazují vyšší reakční dobu stop-signálu (SSRT)při úkolu go / no-go, což odráží horší inhibiční kontrolu, zatímco Forbes et al. (2009) se nepodařilo najít takový účinek na Barrattovu stupnici impulzivity (BIS), nástroj pro měření impulzivity. Na druhou stranu, Colzato et al. (2010) prokázala, že tyto smíšené účinky, může být vzhledem k tomu, že předchozí studie řešit impulzivita jako monolitický proces, zatímco impulzivita může být ve skutečnosti rozdělit na funkční a dysfunkční typy podle Dickman je Impulzivita Zásob (DII; Dickman, 1990). Dysfunkční impulzivita je tendence jednat bez rozmyslu v případech, kdy takové jednání je nevhodné, zatímco funkční impulzivita je podobné tendence realizován ve vhodných situacích (Colzato et al., 2010). Colzato a kol. (2010) zjistili, že jedinci s DRD4 7R, který je spojen s vyššími hladinami striatálního DA, vykazovali vyšší dysfunkční impulzivitu. Několik metaanalytických studií poukázalo na alelu 7R DRD4 jako rizikovou alelu pro poruchu pozornosti s hyperaktivitou (ADHD; Faraone et al. , 2001; DiMaio a kol., 2003), porucha charakterizovaná vysokou dysfunkční impulzivitou (Young et al., 2007). Vztah mezi ADHD a kreativitou byl dále zkoumán v několika studiích (Shaw a Brown, 1990; Healey a Rucklidge, 2006), i když dosud tyto studie přinesly nejasné výsledky. Zatímco někteří zjistili ADHD a jeho symptomatiku být pozitivně korelovala s kreativitou (Healey a Rucklidge, 2006; White a Shah, 2011), jiní uvádějí opačný trend (Funk et al., 1993; Healey and Rucklidge, 2005, 2008). Zajímavé je, že ADHD je behaviorálně spojena s poruchami výkonné kontroly, včetně posunu pozornosti (Boonstra et al ., 2005) a flexibilita (Barkley et al., 1997; seržant a kol., 2002), které jsou důležité pro kognitivní flexibilitu, Ústřední dimenzi tvořivosti (Dietrich, 2004; Durstewitz and Seamans, 2008). Tato zjištění naznačují, že jedinci s alelou 7R DRD4 mohou vykazovat nižší úroveň kreativity a sníženou flexibilitu zejména při úkolech DT. Proto, DRD4 exon III VNTR je biologicky věrohodným kandidátem na přispění k individuálním rozdílům v kreativitě.
kolektivně se zdá, že na jedné straně alela 7R, jako riziková alela pro zhoršené výkonné funkce, flexibilitu a posunutí sady, může být spojena s nízkou kreativitou. Na druhou stranu, asociace mezi alelou 7R a vyšším hledáním novinek naznačuje, že jedinci s alelou 7R mohou ve skutečnosti vykazovat větší flexibilitu a kreativitu.
současná studie byla navržena tak, aby prozkoumala komplexní roli DRD4 7R při částečném formování lidské tvořivosti. Pro lepší charakterizaci tohoto vztahu jsme vyhodnotili dva typy DT testů (figurální a verbální). Bodování jednotlivých úloh byla rozdělena podle tří hlavních dimenzí DT, a sice tvůrčí plynulost, flexibilitu a originalitu, a každé dimenze byl analyzován odděleně testovat hypotézu, že DRD4 rozdílně ovlivňuje každý rozměr kreativity.
metoda
účastníci
vzorek tvořilo 185 studentů z univerzity v Haifě v Izraeli(112 žen). Všichni účastníci byli běloši (sami), s průměrným věkem 24,5 (SD ± 2,1). Účastníci byli přijati prostřednictvím oznámení zveřejněných na univerzitě. Všichni účastníci byli placeni dobrovolníky.
Hodnocení Kreativity
Kreativita byla hodnocena pomocí dvou DT testy, sub-scale, od figurální sub-test Torrance test tvořivého myšlení (Torrance, 1974), a Alternativní Využití Úloh (EL; Guilford et al., 1978). Oba testy zahrnovaly tři základní rozměry DT, a to flexibilitu, originalitu a plynulost.
Torrance Test tvořivého Myšlení (Kruhy Sub-Scale)
Účastníci dostali na stránku, na které 30 identické kruhy byly čerpány. Byli požádáni, aby nakreslili co nejvíce různých smysluplných objektů v časovém limitu 10 min, kde každý výkres musí obsahovat alespoň jeden kruh. Bodování zahrnuty plynulost (počet odpovědí vyrábí), flexibilita (počet kategorií) a originalitu, která se vypočte podle bodování originální odpovědi, jak je uvedeno v Torrance Testy Kreativního Myšlení bodování průvodce (Torrance, 1974).
Alternate Uses Task (AUT)
účastníci dostali seznam pěti běžných objektů a požádali, aby vypsali co nejvíce alternativních použití pro každý objekt v časovém limitu 10 min. Nejběžnější každodenní použití bylo uvedeno v závorce. Předměty byly: bota (běžné použití: opotřebení pěšky); tlačítko (běžné použití: zavírání věcí); tužka (běžné použití: kreslení nebo psaní); pneumatika (běžné použití: kolo automobilu); a sklenice na pití (běžné použití: obsahuje tekutinu). Byly započítány a zahrnuty pouze odpovědi, které neopakovaly dané běžné použití. Stejně jako v testu Torrance zahrnovalo bodování plynulost, flexibilitu a originalitu. Od té doby nejsou žádné pokyny pro bodování v původní odpovědi v AUT. klimatizace, originální odpovědi byly definovány jako statisticky málo časté odpovědi podle předběžné zkoušky prováděné v naší laboratoři.
Pretest
za účelem vytvoření platného kritéria frekvence odezvy dokončila skupina 100 zdravých účastníků, kteří se této studie nezúčastnili, aut. Pro každý objekt byl shromážděn seznam všech možných použití od všech účastníků. Na základě tohoto seznamu bylo vypočteno statistické měření frekvence, aby bylo možné vyhodnotit skóre originality pro každou odpověď, a, následně, pro každého účastníka. Odpovědi bylo přiřazeno skóre nula, jestli 5% nebo více účastníků uvedených na dané použití, skóre jeden, pokud mezi 2% a 4,99% účastníků uvádí, a skóre dva, pokud méně než 1.99% uvedené použití. Průměrné skóre originality bylo vypočteno pro každého účastníka podle těchto statistických skóre frekvence.
extrakce DNA a genotypizace
DNA byla extrahována z 20 ml vzorků ústní vody pomocí Master Pure kit (Epicentre, Madison, WI). DRD4 48-bp VNTR byl charakterizován PCR amplifikace postup s následující primery: F5′ – CTT ACC CCT CTG CCC GCT KOČKA GCT GCTGCT CTA CTGG – 3′ a R5′ – ACC ACCACC GGC AGG ACC CTC ATG GCC TTG CGC TC – 3. PCR reakce byly prováděny za použití 5 µl Master Mix( Thermo scientific), 2 µl primerů (0,5 µM), 0,6 µl Mg/Cl2 (2,5 mM), 0.4 µl DMSO 5% a 1 µl vody na celkový objem 9 µl a do směsi byl přidán další 1 µl genomové DNA. Všechny PCR reakce byly provedeny na termocykleru Biometra T1 (Biometra, Güttingem, Německo). PCR reakce byl stav následující: předehřívání krok v 94.0°C po dobu 5 min, 34 cyklů denaturace při 94.0°C po dobu 30 s, reannealing na 55°C po dobu 30 s, a extenze při 72°C po dobu 90 sekund. Reakce probíhala držet při 72°C po dobu 5 min. Reakční směs byla pak electrophoresed na 3% agarózovém gelu (AMRESCO) s ethidium bromidu na obrazovku pro genotypy.
distribuce genotypové frekvence byla podle Hardy-Weinbergovy rovnováhy (chí-kvadrát = 0,01, hodnota p = 0,92).
Statistické Analýzy
Genotyp byl klasifikovány podle přítomnosti nebo nepřítomnosti 7R alelu genu DRD4 (7 vs. no7), stejně jako v souladu s předchozí zprávy v literatuře (Ding et al., 2002). Skupinové rozdíly ve skóre kreativity byly statisticky analyzovány pomocí ANOVAs, s přítomností (7) nebo nepřítomností (no7) alely DRD4 7R jako nezávislé proměnné. Zkoumat různé aspekty DT (plynulost, flexibilitu, originalitu), jsme provedli multivariační analýzu rozptylu (MANOVA) odděleně pro každou komponentu.
Výsledky
potvrďte, že tyto dvě skupiny (7, no7) se významně nelišily z hlediska věku a vzdělání jsme provedli nezávislé t-testy k porovnání dvou skupin (viz Tabulka 1 průměrů a standardních odchylek). Tato analýza odhalila, že obě skupiny se nelišily z hlediska věku nebo let vzdělání. Neparametrické testy (Mann–Whitney) odhalily významné rozdíly ve frekvenční distribuci pro pohlaví (Z = 2.11, p < 0.05).
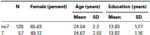
Tabulka 1. Věk, pohlaví a údaje o vzdělání pro genotyp DRD4 7R, SD: směrodatná odchylka.
průměrné skóre opatření kreativity použitých ve dvou úkolech je shrnuto v tabulce 2.

Tabulka 2. Prostředky a standardní chyba (SE) dvou použitých kreativních úkolů: AUT a Torrance.
Plynulost Složka
multivariate ANOVA je uvedeno obecné účinek DRD4 7R genotyp pro plynulost součásti, F(2,182) = 3.25, p < 0.05. Vzhledem k významu celkového testu byly zkoumány jednorozměrné hlavní účinky. Významné jednorozměrné hlavní účinky na genotyp DRD4 byly získány pro AUT plynulost F (1,183) = 4,95, p < 0.05 (7R alela vykazuje nižší plynulost ve srovnání s no7 skupiny, viz Obrázek 1A), ale nedosáhly úrovně významnosti pro Torrance plynulost F(1,183) = 2.95, p = 0.087.
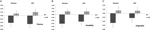
Obrázek 1. Kreativita boduje na úkolech AUT a Torrance oddělených od plynulosti (A), flexibility (B) a originality (C). Prostředky byly přeměněny na skóre Z.
komponenta Flexibility
multivariační ANOVA opatření flexibility vykazovala obecný účinek pro DRD4 7R genotyp F (2,182) = 4.38, p < 0.05. Významné jednorozměrných hlavní efekty pro DRD4 genotypu byly získány pro AUT. klimatizace, flexibilitu, F(1,183) = 4.57, p < 0,05 a pro Torrance flexibilita F(1,183) = 6.95, p < 0.01. Jak je znázorněno na obrázku 1B, v obou případech vykazovali jedinci ve skupině alely 7R nižší flexibilitu ve srovnání s jedinci ve skupině no7 (Tabulka 2).
Originalita Složka
Multivariate ANOVA originality opatření nevykazovaly žádný význam účinky, F(2,182) = 2.06, p = 0.13, ačkoli skóre naznačovalo nižší originalitu pro alelu 7R DRD4 (podrobnosti viz obrázek 1C, tabulka 2).
Genderové Rozdíly
Diskuse
DT úkoly vyžadují, účastníkům poskytovat více řešení daného problému. V úlohách, které se zde používají, byli účastníci povinni myslet na mnoho alternativních možných použití pro každodenní předměty (v AUT) a kreslit více kreseb obsahujících specifický tvar (kruh) (v úkolu Torrance). Výsledky zde uvedené ukazují, že jedinci s alelou DRD4 7R vykazují nižší kreativitu měřenou DT. Vzhledem k tomu, že alela 7R byla spojena s dysfunkční impulzivitou a špatnou inhibicí (Congdon et al ., 2008; Dreber et al., 2009), skutečnost, že jedinci s alelou 7R vykazují nižší kreativitu, lze vysvětlit jejich neschopností potlačit nebo inhibovat zjevné (běžné) reakce, jak naznačuje nižší skóre originality. Nižší skóre flexibility a plynulosti může naznačovat, že jakmile je poskytnuta společná odpověď, je těžší přejít na novou kategorii (jak ukazuje skóre nízké flexibility), což vede k menšímu počtu odpovědí(jak ukazuje skóre nízké plynulosti). Další důkazy o vztahu mezi kreativitou a impulzivitou lze vidět ve spojení alely 7R s ADHD (Li et al ., 2006), stejně jako se zvýšenou impulzivitou u pacientů s ADHD (Langley et al ., 2004). Zajímavé je, že kreativita byla spojena se systémem DA v případě schizofrenie (Eysenck, 1993). Ačkoli několik studií ukazuje, že pacienti se schizofrenií a schizotypální osobností vykazují vysokou kreativitu (O ‚ Reilly et al ., 2001; Folleyovou a Park, 2005), poslední modely schizofrenie a kreativitu podpora obráceného tvaru U sdružení, v němž kreativita může být vyšší, s low-středně schizotypal vlastnost, ale snižuje závažnost psychopatologie u schizofrenie se zvyšuje (Tsakanikos a Claridge, 2005; Stoneham a Coughtrey, 2009; Nelson a Rawlings, 2010). Ve skutečnosti, v nedávné studii Jaracz et al. (2012) uvádí, že pacienti se schizofrenií vykazují sníženou kreativitu ve srovnání s kontrolami. Dále autoři uvádí, že poruchou kreativita je spojena s nízkou kognitivní flexibilitu a impulzivity, odráží v jejich nízké skóre na Wisconsin Card Sorting Test (WCST). Tyto výsledky jsou v souladu s předchozími výzkumy ukazují, že pacienti s chronickou schizofrenií vykazují nižší kreativitu, což v případě kreativní plynulost byla zprostředkována skóre na výkonné řídící úkoly a v případě, že originalita není (Abraham et al., 2007). Kolektivně, zatímco mírné funkční impulzivita může zlepšit kreativitu u pacientů s schizotypal vlastnosti, a to zejména v generování nápadů, které se liší od příkladů uvedených (Abraham a Windmann, 2008), dysfunkční impulzivitu pozorován u těžké schizofrenie může tlumit tvůrčí schopnosti.
výsledky současné studie podporují vztah mezi kreativitou a variantami genetických polymorfismů DA (Reuter et al., 2006; Runco a kol., 2011). Výsledky nalezené v našem vzorku navíc naznačují, že hlavní účinek alely DRD4-7R na kreativitu může být způsoben jejím dopadem na flexibilitu. Několik studií spojilo kognitivní flexibilitu s cestami DA (Ashby a Isen, 1999; Dreisbach a Goschke, 2004; Müller et al., 2007). Bylo postulováno, že aktivace D2-like receptory (tj., D2, D3 a D4) snižuje Gabaergní inhibice a usnadňuje aktivaci více zastoupení, čímž se zvyšuje pružnost se projevuje tím, že set-shifting schopností (Müller et al., 2007; Seamans and Robbins, 2010). Kromě toho byly receptory podobné D2 spojeny s flexibilní integrací nových informací (Durstewitz and Seamans, 2008). V souladu s tím Reuter et al. (2006) našel souvislost mezi dopaminovým receptorem D2 a kreativitou měřenou složeným indexem, ačkoli to, zda flexibilita hrála v těchto výsledcích hlavní roli, nelze určit. Dodatečně, Runco et al. (2011) hlášeny rozdíly v tvůrčí plynulosti mezi nosiči několika dopaminových genů, mezi nimi DRD4. Z jejich zprávy však není jasné, která skupina vykazuje větší tvůrčí schopnosti. Je důležité si uvědomit, že na rozdíl od zde prezentovaných výsledků, Runco et al. (2011) nenašel souvislost mezi opatřeními pružnosti měřenými úkoly DT a DRD4. Tuto nekonzistenci lze vysvětlit rozdíly v použitých populacích a velikostech skupin nebo rozdíly v frekvencích alel v populaci vzorkované. Kromě toho existují důkazy, což naznačuje, že souvislost mezi dopaminem a DT nemusí být lineární (Akbari Chermahini a Hommel, 2010, 2012), což naznačuje, že interakce mezi několika DA geny mohou ovlivnit jednotlivé výsledky každého genetické variability v rámci DT. Kromě toho několik studií poukázalo na skutečnost, že cesty DA mohou být citlivé na pohlaví (Tammimäki a Männistö, 2011; Wang et al., 2012). Zhong et al. (2010) například zjistil, že pohlaví modulovalo souvislost mezi polymorfismem DRD4 exon3 a smyslem pro spravedlnost. V současné studii jsme nenašli interakční účinek mezi DRD4-7R a pohlavím, což naznačuje, že pohlaví nemá modulační účinek na souvislost mezi DRD4 a kreativitou v našem vzorku. Výsledky zde uvedené navíc naznačují nedostatek genderových rozdílů v kreativitě, a zejména v DT (pro přehled viz Baer and Kaufman, 2008).
závěrem v současné studii navrhujeme, že souvislost mezi DRD4 a kreativitou je ovlivněna hlavně flexibilitou. Výsledky aktuální studie jsou v souladu s předchozími výsledky ukazující na zapojení DA systém, v tvořivosti a přidat k hromadění poznatků naznačuje, že genetické vlivy na tvořivost.
existuje několik omezení studie, která by měla být uznána. Za prvé, současná studie zkoumala pouze souvislost mezi jednou genetickou variabilitou v systému DA-DRD4 – a DT. Existují však další důležité polymorfismy ve striatálním DA systému, které by mohly být spojeny s DT a mohou přispívat jak samostatně, tak i v interakcích Gen-gen. Jedním takovým polymorfismem je transportér dopaminu (DAT1), u kterého bylo prokázáno, že je spojen s inhibicí návratu (Colzato et al., 2010) a také s mírou impulzivity (Gizer a Waldman, 2012). Dále, jak bylo uvedeno výše, další možností, která zde nebyla prozkoumána, jsou možné účinky interakce Gen-gen na DT. Akbari Chermahini a Hommel (2010) uvádí, nelineární souvislost mezi značkou dopaminových a DT, což naznačuje, že vztah může být složitější, jako je, že jeden polymorfismus genu mohou mít dopad DT prostřednictvím další genový polymorfismus. Budoucí studie by tedy měly zvážit zkoumání interakce mezi těmito dvěma polymorfismy v kontextu flexibility a DT.
Prohlášení o střetu zájmů
autoři prohlašují, že výzkum byl proveden bez jakýchkoli obchodních nebo finančních vztahů, které by mohly být vykládány jako potenciální střet zájmů.
Akbari Chermahini, s., and Hommel, B. (2010). (B) souvislost mezi kreativitou a dopaminu: spontánní mrknutí sazby předvídat a oddělit divergentní a konvergentní myšlení. Poznání 115, 458-465. doi: 10.1016 / j. poznání.2010.03.007
Pubmed Abstrakt | Pubmed Plný Text / CrossRef Plný Text
ochlazuje, R. (2008). Role dopaminu v motivační a kognitivní kontrole chování. Neurovědec 14, 381-395. doi: 10.1177 / 1073858408317009
Pubmed Abstract | Pubmed Full Text / CrossRef Full Text
Frank, M.J. (2005). Dynamická dopaminová modulace v bazálních gangliích: neurokomputační účet kognitivních deficitů v medikovaném a nemedikovaném parkinsonismu. J. Cogn. Neurovědci. 17, 51–72. doi: 10.1162/0898929052880093
Pubmed Abstraktní | Pubmed Plný Text | CrossRef Úplné Znění
Healey, D., a Rucklidge, j. J. (2006). Zkoumání vztahu mezi symptomatologií ADHD, kreativitou a neuropsychologickým fungováním u dětí. Dětský Neuropsychol. 12, 421–438. doi: 10.1080/09297040600806086
Pubmed Abstraktní | Pubmed Plný Text | CrossRef Úplné Znění
Runco, M. a., a Richards, R. (1997). Vynikající kreativita, každodenní kreativita a zdraví. Greenwich, ČT: Ablex.
Torrance, e. (1974). Rozdíly nejsou deficity. Učit. SB. Záznam. 75, 471–487.