- Úvod
- Materiály a Metody
- Rostlinné Materiály a Růstu Stavu
- PSI a PSII Měření
- Elektrochromní Shift Analýzy
- Fotoinhibiční ošetření
- Statistická analýza
- Výsledky
- diskuse
- Role Stimulace CEF při slabém osvětlení
- Změna oxidačního poměru P700 při slabém osvětlení nesouvisí s pmf
- ETRII Kontroly P700 Oxidace Poměr při Nízké Světlo
- závěr
- Autor Příspěvky
- finanční Prostředky
- Prohlášení o střetu zájmů
Úvod
i když světlo je hybnou silou fotosyntézy, vysoká světlo může způsobit značné photoinhibition z fotosystému II (PSII) pro listy odstín-založení rostlin (Kitao et al., 2000; Barth a kol., 2001; Krause a kol., 2004; Huang et al., 2015b, 2016b, c). Fotoinhibovaná PSII se však rychle zotavuje při slabém osvětlení (He a Chow, 2003; Zhang and Scheller, 2004, Allakhverdiev et al., 2005), kvůli rychlému obratu proteinu D1 (Aro et al., 1993; Zhang a Scheller, 2004; Allakhverdiev et al., 2005). Rychlost opravy PSII byla snížena po inhibici syntézy ATP buď pomocí PSI nebo PSII (Allakhverdiev et al ., 2005), což naznačuje, že rychlá oprava fotodynamicky poškozených komplexů PSII potřebuje v krátké době velké množství ATP. Předchozí studie ukázala, že po ochlazení indukované fotoinhibici PSII byl cyklický tok elektronů (CEF) významně stimulován během zotavení při slabém osvětlení (Huang et al., 2010). Bylo navrženo, že tato stimulace CEF při slabém osvětlení zvýšila syntézu ATP pro rychlou opravu PSII (Huang et al., 2010). K objasnění specifické úlohy stimulace CEF při slabém osvětlení po fotoinhibici PSII je však zapotřebí více důkazů.
během CEF se elektrony z NADPH nebo ferredoxinu cyklují kolem PSI do plastochinonového fondu. Tento přenos elektronů je spojen se protonové translokace a vytváří protonový gradient přes membrány thylakoidu (ΔpH) (Shikanai a Yamamoto, 2017). Kromě přispění syntézy ATP, další funkce ΔpH je down-regulaci fotosyntetických elektronový transport po okyselení lumen thylakoidu (Shikanai, 2014, 2016). Toto nařízení zahrnuje dva různé mechanismy: jedna se týká tepelné energie, tepla a odvádí přebytečné vstřebává světelnou energii jako teplo z antény PSII (Takahashi et al., 2009) a druhá je down-regulace komplexní aktivity Cyt b6/f a řídí rychlost přenosu elektronů na PSI (Suorsa et al ., 2012, 2016; Tikkanen a Aro, 2014). V angiospermech včetně Arabidopsis thaliana fungují dvě cesty cyklického elektronového transportu PSI (Shikanai, 2007). První CEF dráha je pgr5-/PGRL1-dependentní dráha citlivá na antimycin A (Munekage et al ., 2002; Sugimoto a kol., 2013) a druhý je závislý na komplexu podobném chloroplastu NADH (NDH) (Burrows et al., 1998; Shikanai et al., 1998). Příspěvek dráhy závislé na PGR5- / PGRL1 je významnější u rostlin C3. Za vysokého světla aktivace CEF závislého na PGR5-/PGRL1 indukuje okyselení tylakoidního lumenu a vede tak k vysokým hladinám oxidačního poměru P700 (Suorsa et al ., 2012, 2016; Kono a kol., 2014; Yamori a kol., 2016). Pro srovnání, oxidační poměr P700 při slabém osvětlení byl málo ovlivněn nedostatkem CEF závislého na PGR5-/PGRL1 (Munekage et al ., 2002, 2004; Kono a kol., 2014). Při mírné fotoinhibici PSII byla stimulace CEF při slabém osvětlení doprovázena zvýšením oxidační dávky P700 (Huang et al ., 2010). Není však jasné, zda je tento zvýšený oxidační poměr P700 způsoben stimulací CEF.
kromě CEF hraje přenos elektronů z PSII (ETRII) důležitou roli při ovlivňování redoxního stavu PSI (Tikkanen et al., 2014; Huang et al., 2016a, c; Suorsa a kol., 2016). V pgr5 rostliny A. thaliana, závažné photoinhibition z PSII je pravděpodobné, že funkce jako maximální kontrolu nad fotosyntetického přenosu elektronů umožňující údržbu P700 optimálně oxiduje za přebytečné světlo (Tikkanen et al., 2014). Nedávno, Suorsa a kol. (2016) ukázalo, že rostliny pgr5 vykazovaly vysoký oxidační poměr P700 za vysokého světla, když byla etrii snížena kombinací vyřazovacích mutací v lokusech PsbO1, PsbP2, PsbQ1, PsbQ2 a PsbR. V chlazených listech tabáku mírná fotoinhibice PSII stlačila tok elektronů na PSI a poté zvýšila oxidační poměr P700 během dalšího chlazení (Huang et al ., 2016a). Kromě toho chladem indukovaná fotoinhibice PSII vedla k depresi ETRII, která byla doprovázena zvýšením oxidačního poměru P700 při slabém osvětlení (Huang et al., 2010). Předpokládáme, že oxidační poměr P700 při slabém osvětlení je primárně řízen proudem elektronů z PSII.
V naší předchozí zprávě, pozorovali jsme, že odstín, kterým se rostlina Panax notoginseng ukázal selektivní photoinhibition z PSII pod high-light stresu. Po krátkodobém vystavení vysokému světelnému stresu se ETRII při slabém osvětlení významně snížil, ale CEF byl významně stimulován. Současně se oxidační poměr P700 do značné míry zvýšil. Naším specifickým cílem bylo (1) prozkoumat, zda stimulace CEF při slabém osvětlení usnadňuje hlavně syntézu ATP; a (2) Určete, zda zvýšení oxidačního poměru P700 při mírné fotoinhibici PSII souvisí spíše s ETRII než se stimulací CEF nebo se změnou pmf. Za účelem řešení těchto otázek byly neporušené listy Panax notoginsengu ošetřeny při 2258 µmol fotony m-2 s-1 po dobu 30 minut. Před a po ošetření vysokým světlem byla stanovena distribuce energie v PSII, redoxní stav PSI a protonová hnací síla (pmf) při slabém světle 54 µmol fotonů m-2 s-1.
Materiály a Metody
Rostlinné Materiály a Růstu Stavu
V této studii, 2-roky-starý rostlin odstín-založení rostlina Panax notoginseng (Burkill) F. H. Chen ex C. Chow a. W. G. Huang byly použity pro experimenty. Tyto rostliny byly pěstovány při světelném stavu 10% slunečního světla (s maximální intenzitou denního světla přibližně 200 µmol fotonů m-2 s-1). U těchto rostlin nebyly zaznamenány žádné stresy vody a výživy. Pro fotosyntetická měření byly použity 9týdenní plně rozšířené listy.
PSI a PSII Měření
Fotosystému i a PSII parametry byly měřeny při teplotě 25°C tím, že současně zaznamenaných pomocí Dual PAM-100 měřicí systém (Heinz Walz GmbH, Effeltrich, Německo). Fluorescenční parametry chlorofylu byly vypočteny následovně: Fv / Fm = (Fm-Fo) / Fm, Y (II) = (Fm ‚- Fs) / Fm‘ (Genty et al., 1989), NPQ = (Fm-Fm‘) / Fm‘. Fo a Fm jsou minimální a maximální fluorescence po tmavé aklimatizaci. Fs je světelně přizpůsobená fluorescence v ustáleném stavu. Fo a Fm byly stanoveny po aklimatizaci za tmy po dobu 30 minut před a po ošetření vysokým světlem. PSI fotosyntetické parametry byly měřeny pomocí Dual PAM 100 na základě P700 signál (rozdíl intenzit 830 a 875 nm puls-modulované měření světla dopadajícího na fotodetektor) (Klughammer a Schreiber, 2008). Signály P700+ (P) se mohou lišit mezi minimální (P700 plně redukovanou) a maximální hladinou (P700 plně oxidovaný). Maximální úroveň (Pm) byla stanovena za použití saturační pulz (600 ms a 10000 µmol fotonů m-2 s-1) po pre-osvětlení s far-red světlo, a Odpoledne byl použit pro odhad PSI aktivity. Pm ‚ byl stanoven podobně jako Pm, ale s aktinickým světlem místo daleko červeného světla. Kvantový výtěžek PSI byl vypočítán jako Y (I) = (Pm‘ – P) / Pm. Oxidační poměr P700 v daném aktinickém světle byl vypočítán jako Y (ND) = P/Pm. Kvantový výtěžek PSI nefotochemického rozptylu energie v důsledku omezení na straně akceptoru byl vypočítán jako Y (na) = (Pm-Pm‘)/Pm. Hodnoty ustáleného stavu pro Y (II), NPQ, Y(I) a Y(ND) byly měřeny po aklimatizaci světla při slabém osvětlení po dobu 20 minut.
Fotosyntetických tok elektronů přes PSI a PSII byly vypočteny jako: ETRII = Y (II) ×PPFD ×0.84 ×0.5 (Krall a Edwards, 1992), ETRI = Y (I) ×PPFD×0.84 ×0.5 (Yamori et al., 2011), kde se předpokládá, že 0,5 je podíl absorbovaného světla dosahujícího PSI nebo PSII, a 0,84 je absorbance (zlomek dopadajícího světla absorbovaného listy). Rozsah aktivace CEF byl odhadnut jako poměr ETRI/ETRII (Yamori et al., 2011, 2015). Je třeba poznamenat, že rozdělení excitace 1:1 mezi PSI a PSII nemusí být skutečné, protože stínové rostliny mají tendenci zvyšovat poměr PSII/PSI (Lunde et al ., 2003; Tikkanen et al., 2006; Grieco a kol., 2012). Dále v této studii byly parametry PSI a PSII měřeny pomocí červeného aktinického světla (635 nm) poskytovaného společností Dual-PAM 100 (Walz, Německo). Protože červené aktinické světlo upřednostňuje excitaci PSII před excitací PSI, může být distribuce excitační energie z LHCII na PSII a PSI ovlivněna červeným aktinickým světlem (Tikkanen et al ., 2017).
Elektrochromní Shift Analýzy
ECS signálu byla sledována jako změna absorbance při 515 nm za použití Dual-PAM-100 (Walz, Effeltrich, Německo), vybavený P515/535 zářič-detektor modul (Walz). ECS signál byl získán po 20 min. osvětlení na 54 µmol fotonů m-2 s-1, poté, ECS rozpadu byla měřena vypnutí aktinického světla pro 30 sekund. Analýza ECS tmavě interval odpočinku kinetika (DIRKECS) bylo provedeno metodou Sacksteder et al. (2001) a Takizawa et al. (2008). Celkový pmf byl odhadnut z celkové amplitudy rychlého rozpadu signálu ECS během tmavého intervalu 300 ms. Pomalá relaxace signálu ECS umožnila rozpoznat příspěvek protonového gradientu přes tylakoidní membrány (ΔpH). Časová konstanta relaxace ECS prvního řádu (tec) je nepřímo úměrná protonové vodivosti (gH+) tylakoidní membrány prostřednictvím ATP syntázy (Sacksteder a Kramer, 2000; Cruz a kol., 2005). V důsledku toho byl GH+ odhadnut jako inverzní konstanta doby rozpadu .
Fotoinhibiční ošetření
po tmavé aklimatizaci po dobu 30 minut byly měřeny Fv / Fm a Pm v neporušených listech. Poté byly tyto neporušené listy přizpůsobeny světlu při 59 µmol fotonech m-2 s-1 po dobu 20 minut a byly zaznamenány parametry fluorescence chlorofylu, signálu P700 a signálu ECS. Poté bylo aktinické světlo změněno na 2258 µmol fotonů m-2 s-1. Po vystavení tomuto vysokému světlu po dobu 30 minut bylo aktinické světlo okamžitě změněno na 59 µmol fotonů m-2 s-1 a fotosyntetické parametry byly zaznamenány po aklimatizaci světla po dobu 20 minut. Nakonec byly FV / Fm a Pm měřeny po tmavé aklimatizaci po dobu 30 minut.
Statistická analýza
výsledky byly zobrazeny jako průměrné hodnoty pěti nezávislých experimentů. Nezávislý t-test byl použit na úrovni významnosti α = 0,05 k určení, zda existují významné rozdíly mezi různými léčbami. Všechny statistické analýzy byly provedeny pomocí SPSS 16.0.
Výsledky
Během indukce fotosyntézy na 59 µmol fotonů m-2 s-1, ETRII postupně zvyšovala a dosáhla rovnovážného stavu asi 18 min (Obrázek 1A). Pro srovnání, ETRI byl během této indukční fáze konstantní (obrázek 1A). Po nástupu tohoto slabého světla byl ETRI mnohem vyšší než ETRII. Po 20minutové fotosyntetické indukci byl však ETRI nižší než ETRII. Tyto výsledky naznačily, aktivace CEF během počáteční fáze indukce, v souladu s předchozí studií (Joliot a Joliot, 2002, 2005; Makino et al., 2002). Po této fotosyntetické indukci při slabém osvětlení byly listy osvětleny při vysokém světle 2258 µmol fotonů m-2 s-1 po dobu 30 minut. Je zajímavé, že ETRII se během léčby vysokým světlem postupně snižovala (obrázek 1B). Mezitím byla hodnota ETRI vyšší než hodnota ETRII (obrázek 1B), což naznačuje aktivaci CEF za vysokého světla. Po tomto ošetření vysokým světlem byl maximální fotooxidovatelný P700 (Pm) udržován stabilní (obrázek 2A). Pro srovnání, maximální intenzita fluorescence (Fm) se snížila o 40% a maximální kvantový výtěžek PSII (Fv/Fm) se snížila od 0,80 0,65 (Obr. 2B,C). Tyto výsledky ukázaly selektivní fotoinhibici PSII v listech Panax notoginsengu při vysokém světelném stresu.
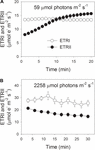
obrázek 1. Fotosyntetický tok elektronů při slabém světle (A) a vysokém světle (B). (A) Tmavě aklimatizovat listy byly vystaveny nízké světlo 59 µmol fotonů m-2 s-1 po dobu 20 min. (B)po osvětlení při 59 µmol fotonech m-2 s-1 po dobu 20 minut pro aktivaci elektronového dřezu ve fotosyntéze byly listy vystaveny vysokému světlu 2258 µmol fotonů m-2 s-1 po dobu 30 minut. Hodnoty jsou prostředky ± se (n = 5).
dříve, Než high-light ošetření zralé listy byly osvětleny v nízkých světelných 59 µmol fotonů m-2 s-1 po dobu 20 min pro aktivaci fotosyntézy, a pak hodnoty pro ETRI, ETRII, Y(ND), a NPQ byly zaznamenány. Po high-light ošetření po dobu 30 min, hodnoty pro ETRI, ETRII, Y(ND), a NPQ byly zaznamenány následující nové aklimatizace fáze 20 min na mírném světle. Před ošetřením vysokým světlem byly hodnoty pro ETRI a ETRII při 59 µmol fotonech m-2 s-1 13,4 a 15,7 µmol elektronů m-2 s-1 (Obrázky 3A, B). Po ošetření vysokým světlem byly etri a ETRII při slabém osvětlení 12,8 a 11,2 µmol elektronů m-2 s-1 (Obrázky 3A, B). Hodnota ETRII při 59 µmol fotonech m-2 s-1 se po fotoinhibiční léčbě snížila přibližně o 30%, což naznačuje depresi ETRII při mírné fotoinhibici PSII. Před léčbou vysokým světlem byla hodnota poměru ETRI/ETRII při 59 µmol fotonech m-2 s-1 0,85 (obrázek 3C). Po fotoinhibiční léčbě se poměr ETRI/ETRII významně zvýšil na 1, 14 (obrázek 3C). Tyto výsledky naznačují stimulaci CEF při slabém osvětlení při fotoinhibici PSII (Yamori et al ., 2011, 2015).
Po high-light ošetření v ustáleném stavu hodnota Y(I) na 59 µmol fotonů m-2 s-1 se nezměnila (Obrázek 4A). Zajímavé je, že Y(NA) výrazně klesl z 0,36 na 0,23 (Obrázek 4B), a Y(ND), se výrazně zvýšil od 0,1 do 0,25 (Obrázek 4C). Tyto výsledky ukázaly změnu redoxního stavu PSI při slabém osvětlení po fotoinhibiční léčbě. Mezitím se hodnota NPQ mírně změnila (obrázek 4D). V důsledku toho měla střední fotoinhibice PSII různé účinky na oxidační poměr P700 a NPQ při slabém osvětlení.
s cílem objasnit, zda zvýšení Y(ND) je způsoben zvýšením pmf, elektrochromní posun signálů na 54 µmol fotonů m-2 s-1 byly stanoveny po světle aklimatizace po dobu 20 min před a po střední-světlo léčba. Je zajímavé, že celkový pmf se významně snížil o 18% po ošetření vysokým světlem (obrázek 5A), ale hladina ΔpH se nepatrně změnila(obrázek 5B). Protože tvorba pmf může být ovlivněna thylakoidu protonové vodivosti, protonové vodivosti (gH+) začaly s buňkou vyvíjet společně na této nízké světlo bylo také zkoumáno, před a po střední-světlo léčba. Výsledek ukázal, že gH+ nezměnil s high-light ošetření (Obrázek 5C), což naznačuje, že high-light ošetření stěží vliv na aktivitu chloroplastů ATP syntázy v nízké světla. Protože pokles o 30% v ETRII byl doprovázen poklesem pmf o 18%, stimulace CEF při slabém osvětlení částečně kompenzovala tvorbu pmf a ΔpH.

obrázek 5. Hodnoty pro celkové proton motive force (pmf) (A), protonový gradient (ΔpH) přes membrány thylakoidu (B), a protonová vodivost (gH+) v membráně thylakoidu (C) v nízké světla. Všechny parametry byly měřeny po adaptaci na 54 µmol fotonů m-2 s-1 po dobu 20 minut. Hodnoty jsou prostředky ± se (n = 5). Hvězdička označuje významnou změnu (P < 0,05) po ošetření vysokým světlem.
diskuse
Role Stimulace CEF při slabém osvětlení
bylo uvedeno, že CEF hraje důležitou roli při udržování fotosyntézy a růstu rostlin (Yamori a Shikanai, 2016). Za vysokého světla se CEF nyní považuje za nezbytný pro vyvážení energetického rozpočtu ATP / NADPH a pro ochranu PSI a PSII před poškozením (Munekage et al ., 2002, 2004; Takahashi et al., 2009; Suorsa a kol., 2012, 2016; Walker a kol., 2014; Huang et al., 2015a, 2017b). Při nízké intenzitě světla hraje CEF důležitou roli při optimalizaci fotosyntetické asimilace CO2 pravděpodobně prostřednictvím dodávky extra ATP (Yamori et al ., 2011, 2015; Nishikawa et al., 2012; Huang et al., 2015a). Hlavní role CEF je pružně modulována v reakci na intenzitu světla. Zejména nástroj pro propojení evropy především přispívá k vyrovnávání ATP/NADPH energii rozpočtu v rámci sub-nasycení svítivosti, ale hlavně chrání fotosyntetického aparátu proti photoinhibition, když je vystavena nasycení svítivosti prostřednictvím acidifikace lumen thylakoidu (Huang et al., 2015a). Zajímavě, Huang et al. (2010) zjistil, že CEF byl významně stimulován při slabém osvětlení po fotoinhibici PSII vyvolané chlazením a předpokládal, že tato stimulace CEF hlavně zvýšila syntézu ATP pro rychlou opravu PSII. K podpoře této hypotézy však bylo zapotřebí více důkazů. V této studii jsme pozorovali, že hodnota etri / ETRII při slabém osvětlení se významně zvýšila (obrázek 3C), což naznačuje stimulaci CEF kolem PSI (Huang et al ., 2011, 2012, 2017a, b; Yamori a kol., 2011, 2015). Současně se amplituda pmf významně snížila a ΔpH mírně poklesla. Tyto výsledky ukázaly, že stimulace CEF neměl vyvolat ΔpH-závislé down-regulaci fotosyntetického transportu elektronů. Jinými slovy, tato stimulace CEF při slabém osvětlení hlavně usnadnila syntézu ATP.
i když protonové vodivosti chloroplastu ATP syntázy nebyla ovlivněna high-light ošetření, menší velikost pmf mělo za následek snížení skutečné rychlosti syntézy ATP. Rychlosti opravy PSII a fotosyntetické asimilace CO2 při slabém osvětlení jsou omezeny hlavně rychlostí syntézy ATP (Allakhverdiev et al., 2005; Yamori a kol., 2011, 2015; Nishikawa et al., 2012). Protože etrii významně poklesla při fotoinhibici PSII, rychlost syntézy ATP pomocí ETRII byla pozoruhodně snížena. Aby bylo možné vyvážit poměr ATP / NADPH vyžadovaný primárním metabolismem a rychlou opravou PSII, je nutný flexibilní mechanismus k zajištění extra ATP. Za této podmínky se ETRII snížil o 30%, ale celkový pmf se snížil pouze o 18%. Tyto výsledky naznačují, že stimulace CEF kompenzovat snížení ETRII závislé na tvorbě pmf, zvýšení syntézy ATP. Rychlá oprava fotodamaged PSII je závislá na syntéze ATP (Allakhverdiev et al., 2005). Zrychlení CEF kolem PSI zvýšilo intracelulární koncentraci ATP, čímž urychlilo rychlost opravy PSII u Synechocystis (Allakhverdiev et al ., 2005). Společně navrhujeme, že stimulace CEF při slabém osvětlení hraje důležitou roli v rychlé opravě aktivity PSII prostřednictvím další syntézy ATP.
Změna oxidačního poměru P700 při slabém osvětlení nesouvisí s pmf
zajímavým jevem je, že oxidační poměr P700 při slabém osvětlení se významně zvýšil při mírné fotoinhibici PSII (Huang et al ., 2010), což je také ukázáno v této studii. Za podmínek, kdy absorbované světlo překračuje požadavky na fotosyntézu, generace ΔpH závislá na CEF reguluje aktivitu komplexní aktivity Cyt b6/f a řídí tok elektronů z PSII do PSI (Suorsa et al ., 2012, 2016; Shikanai, 2014, 2016), optimalizace redoxního stavu P700 v PSI a minimalizace produkce ROS v PSI během fotosyntézy. V pgr5-rostlinách a. thaliana je P700 redukován elektrony ve světle kvůli ztrátě generace pmf. Nicméně, v pgr5 mutant hromadí flavodiiron bílkoviny, úroveň pmf byla také obnovena wild-type úrovni, a tak P700 bylo oxidováno jako ve wild-type (Yamamoto et al., 2016). V liniích mutantů pšenice s deficitem chlorofylu b vede nedostatečný gradient tylakoidních protonů k nadměrné redukci strany akceptoru PSI a tím k FOTOINHIBICI PSI za vysokého světla nebo vysoké teploty (Brestic et al., 2015, 2016). V poslední době, některé studie uvádí, že zvýšená aktivita chloroplastů ATP syntázy poruchou tvorby pmf a způsobil více než-snížení fotosyntetického elektronového transportního řetězce, což vede k fotopoškození PSI pod světlo vysoké a kolísavé světlo (Kanazawa et al., 2017; Takagi a kol., 2017). Tyto zprávy podporují kritickou úlohu pmf při udržování optimálně oxidovaného P700 za nadměrné světelné energie.
Naše současné výsledky ukazují, že po high-light ošetření po dobu 30 min, celkový pmf tvořil v tomto slabém světle výrazně poklesly a tvorbu ΔpH mírně změnila (Obrázky 5A,B). Mezitím se oxidační poměr P700 významně zvýšil z 0,1 (před ošetřením) na 0,25 (po ošetření) (obrázek 4C). Tyto výsledky silně naznačují, že toto zvýšení oxidačního poměru P700 nelze vysvětlit změnou pmf, která se do značné míry liší od korelace mezi Y(ND) a pmf za vysokého světla (Yamamoto et al ., 2016; Takagi a kol., 2017). Proto je regulační účinek pmf na oxidační poměr P700 minimální při slabém osvětlení, ale stává se obzvláště důležitým za vysokého světla nebo kolísajícího světla. Za vysokého světla je vysoká hodnota Y (ND) obvykle doprovázena vysokou hodnotou NPQ (Munekage et al., 2002, 2004; Kono a kol., 2014; Živčák a kol., 2015), kvůli nárůstu pmf. Zhoršení tvorby ΔpH vede ke snížení hladin Y (ND) a NPQ za vysokého světla (Suorsa et al., 2012; Kono a kol., 2014; Yamamoto a kol., 2016; Kanazawa a kol., 2017). Indukce NPQ při slabém osvětlení je určena hlavně úrovní okyselení lumenu. Po ošetření vysokým světlem se rozsah okyselení lumenů nezměnil (obrázek 5B) a NPQ zůstal stabilní (obrázek 4D). Pro srovnání, Y (ND) výrazně vzrostl. Tyto výsledky ukazují, že při slabém osvětlení byly Y (ND) a NPQ řízeny různými regulačními mechanismy.
ETRII Kontroly P700 Oxidace Poměr při Nízké Světlo
PSI se stává over-snížena pouze tehdy, když tok elektronů od PSII přesahuje kapacitu PSI elektronové akceptory vyrovnat se s elektrony. Když je tok elektronů do PSI omezen, PSI je extrémně tolerantní vůči světelnému stresu v rostlinách pgr5 a. thaliana (Suorsa et al ., 2012, 2016; Tikkanen et al., 2014). Pro odstín-založení rostliny Psychotria rubra, PSI činnosti byl odolný vůči high-light stresu v přítomnosti DCMU (Huang et al., 2016c). V chlazených listech tabáku umožnila mírná fotoinhibice PSII udržení optimálně oxidované a poté chráněné aktivity PSI proti dalšímu fotodamáži (Huang et al ., 2016a). Tyto výsledky ukázaly důležitou roli ETRII při kontrole redoxního stavu PSI při vysokém světelném nebo chladivém namáhání. Naše současné výsledky ukázaly, že mírná fotoinhibice PSII vedla k významné depresi ETRII při slabém osvětlení. Mezitím se aktivita PSI a aktivita chloroplastové ATP syntázy mírně změnily a celkový pmf a ΔpH se nezvýšily. Jako výsledek, snížení ETRII nebyla způsobena ΔpH-závislé fotosyntetických ovládání přes Cyt b6/f komplex, ale byl pravděpodobně vyvolán poklesem aktivity PSII (Tikkanen et al., 2014). Snížená dodávka elektronů z PSII na PSI vedla ke zvýšené úrovni oxidace P700. Oxidační poměr P700 při slabém osvětlení byl tedy z velké části řízen ETRII.
závěr
zjistili jsme, že selektivní fotoinhibice PSII vyvolala stimulaci CEF a zvýšení oxidačního poměru P700 při slabém osvětlení. Stimulace CEF nespustila down-regulaci fotosyntetického transportu elektronů závislou na ΔpH. Výsledkem je, že tato stimulace CEF při slabém osvětlení usnadnila hlavně syntézu ATP, která je nezbytná pro rychlou opravu fotodamaged PSII. Zvýšení P700 oxidace poměr nemůže být vysvětlen změnou v ΔpH-závislé fotosyntetických kontroly na Cyt b6/f komplex, ale byl způsoben především sníženou nabídku elektrony od PSII k PSI.
Autor Příspěvky
WH, S-BZ, a TL koncipovaný a navržený výzkum. WH a Y-JY provedli experimenty. WH, Y-JY, S-BZ, a TL analyzovali data. WH napsal první návrh rukopisu, který byl intenzivně editován všemi autory.
finanční Prostředky
Tato studie byla podpořena Národní Přírodní Science Foundation Číny (31670343), Mládeže Inovace a Propagace Asociace Čínské Akademie Věd (2016347), a Major Vědy a Techniky Programy v Provincii Yunnan (2016ZF001).
Prohlášení o střetu zájmů
autoři prohlašují, že výzkum byl proveden bez jakýchkoli obchodních nebo finančních vztahů, které by mohly být vykládány jako potenciální střet zájmů.
Kono, m., Noguchi, K., and Terashima, i. (2014). Role cyklického toku elektronů kolem PSI (CEF-PSI) a alternativních drah závislých na O2 v regulaci fotosyntetického toku elektronů v krátkodobém kolísavém světle v Arabidopsis thaliana. Fyziol Rostlinných Buněk. 55, 990–1004. doi: 10.1093/pcp/pcu033
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Shikanai, T. (2016). Regulační síť protonové hnací síly: příspěvek cyklického transportu elektronů kolem fotosystému i. Fotosynth. Rez.129, 253-260. doi: 10.1007/s11120-016-0227-0
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Yamori, W., Makino, A., a Shikanai, T. (2016). Fyziologická úloha cyklického přenosu elektronů kolem fotosystému I při udržování fotosyntézy za kolísajícího světla v rýži. Věda. Rep. 6: 20147. doi: 10.1038/srep20147
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Yamori, W., a Shikanai, T. (2016). Fyziologické funkce cyklického transportu elektronů kolem fotosystému I při udržování fotosyntézy a růstu rostlin. Annu. Rev Plant Biol. 67, 81–106. doi: 10.1146/annurev-arplant-043015-112002
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Yamori, W., Shikanai, T., a Makino, A. (2015). Fotosystém i cyklický tok elektronů přes komplex podobný chloroplastu NADH dehydrogenázy plní fyziologickou roli při fotosyntéze při slabém osvětlení. Věda. Rep. 5: 13908. doi: 10.1038/srep13908
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Zhang, S. P., a Scheller, H. V. (2004). Fotoinhibice fotosystému i při chladné teplotě a následné zotavení u Arabidopsis thaliana. Fyziol Rostlinných Buněk. 45, 1595–1602. doi: 10.1093/pcp / pch180
PubMed Abstrakt / CrossRef Plný Text / Google Scholar