- Introduction
- menetelmä
- osallistujat
- luovuuden arviointi
- Torrance Test of Creative Thinking (Circles Sub-Scale)
- Alternative Uses Task (AUT)
- Pretest
- DNA: n uuttaminen ja genotyypin määritys
- tilastollinen analyysi
- tulokset
- Sujuvuuskomponentti
- Joustavuuskomponentti
- alkuperäisyyskomponentilla
- sukupuolierot
- Keskustelu
- Eturistiriitalausunto
Introduction
Creative cognition is a important role in the arts, in invention and innovation, as well in everyday life (Runco and Richards, 1997). Vaikka luovuutta on pidetty ainutlaatuisena ihmisen kykynä, spontaanisti luovaa käyttäytymistä (esim., creating new tools among primates and birds) on osoitettu tapahtuvan myös muilla kuin ihmisillä (Byrne and Bates, 2007), mikä todistaa luovuuden syvistä evolutionaarisista ja biologisista juurista. Luovuus on määritelty kyvyksi tuottaa vastauksia, jotka ovat sekä uusia (eli alkuperäisiä, harvinaisia ja odottamattomia) että sopivia (eli mukautuvia ja hyödyllisiä tehtävärajoitusten mukaan) (Sternberg and Lubart, 1999). Yksi psykometrisistä lähestymistavoista luovuuden mittaamiseen liittyy divergent thinking (DT) – tehtäviin, joissa osallistujia pyydetään vastaamaan tiettyyn ongelmaan useilla ratkaisuilla (Dietrich and Kanso, 2010). DT: n testit mittaavat yleensä luovuuden eri osa-alueita, kuten luovaa sujuvuutta, joustavuutta ja omaperäisyyttä (Torrance, 1974). Vaikka DT-tehtävät eivät siis olekaan synonyymejä luovuudelle, ne tarjoavat jäsenneltyjä ja objektiivisia luovuuden mittauksia (Sternberg and Lubart, 1999; Jung et al., 2009). Erityisesti DT-tehtävien pistemäärien on osoitettu korreloivan positiivisesti ekologisesti pätevien luovien saavutusten mittareiden kanssa (Carson et al., 2005) sekä itsearvostellulla luovuudella (Furnham and Bachiar, 2008).
aiemmat tutkimukset ovat osoittaneet dopaminergisen (DA) järjestelmän osallisuuden luovuuteen (Heilman ym., 2003; Flaherty, 2005; Takeuchi et al., 2010). Takeuchi ym. (2010) havaitsi, että yksilölliset erot luovuudessa, mitattuna DT: llä, korreloivat positiivisesti harmaaseen aineeseen da-järjestelmän alueilla, mukaan lukien dorsolateraalinen etuotsalohko, bilateraalinen tyvitumake, substantia nigra ja tegmentaalinen ventraalinen alue. Lisäksi useat geneettiset tutkimukset ovat osoittaneet DT: n ja dopamiinin neurotransmission välisen suhteen (Reuter et al., 2006; De Manzano et al., 2010; Runco ym., 2011). Esimerkiksi Reuter et al. (2006) havaitsi, että luovuus, mitattuna DT-tehtävillä, joihin liittyy sekä kuvaannollista että verbaalista luovuutta, liittyy merkittävästi dopamiini-D2-reseptorigeenin (DRD2) polymorfismeihin. Lisäksi useissa tutkimuksissa on havaittu aivokuoren dopamiinin osallistuvan kognitiiviseen joustavuuteen (Frank, 2005; Cools, 2008; Garcia-Garcia et al., 2010), yksi DT: n pääkomponenteista.
yksi kiinnostava DA-kandidaattigeeni luovuudelle on dopamiini D4-reseptorigeeni (DRD4). DRD4-reseptori on yksi viidestä dopamiinireseptorista ja sillä on tärkeä rooli synaptisten dopamiinisignaalien välittäjänä. Geenille on ominaista 48 emäsparin muuttuva määrä tandem-toistoja (48-bp VNTR), jotka sijaitsevat kolmannen eksonin koodausalueella. DRD4 48-bp VNTR polymorfismi vaihtelee 2-11 toistoa yksilöiden välillä (Asghari et al., 1995; Cravchik and Goldman, 2000), jossa 4-toisto (4R) on yleisin toisto Kaukasialaisissa populaatioissa ja 7-toisto (7R) on toiseksi yleisin muunnos (Chang et al., 1996; Ding et al., 2002). Mielenkiintoista, 7R on aiemmin liitetty tosielämän luovaa käyttäytymistä, kuten uutuus-etsivät persoonallisuuden piirre (Ebstein et al., 1996). Uutuudenhakua—taipumusta tutkivaan toimintaan-pidetäänkin yhtenä luovien ihmisten ominaispiirteistä (Chavez-Eakle et al., 2006; Drago et al., 2009). Vaikka todisteet DRD4 7R: n ja uutuuden etsimisen välisestä yhteydestä ovat epäjohdonmukaisia, kuten useat meta-analyysiraportit osoittavat (Kluger et al., 2002; Munafò et al., 2008), monet tutkimukset ovat löytäneet merkittävän yhteyden uutuuden etsimisen ja DRD4: n 7R-alleelin välillä (Ebstein et al., 1996; Benjamin et al., 2000; Keltikangas-Järvinen ym., 2002; Becker ym., 2005). Roussos ym. (2009) ovat äskettäin ehdottaneet, että erot uutuuden etsimisen mittaamisessa omailmoitusasteikoilla voivat selittää epäjohdonmukaiset tulokset 7R polymorfismin ja uutuuden etsimisen välillä. Tämän mukaisesti on toistuvasti todettu, että DA-järjestelmällä on suuri rooli uutuudenhakuisuuden persoonallisuuspiirteessä (Flaherty, 2005; Schweizer, 2006), mikä todistaa edelleen DRD4: n mahdollisesta roolista luovuudessa.
kuitenkin, toisin kuin DRD4 48-bp VNTR 7R: n rooli uutuuden etsimisessä, viimeaikaiset todisteet osoittavat, että 7R todella liittyy heikentyneeseen kognitiiviseen joustavuuteen, joka on yksi luovuuden tärkeimmistä näkökohdista (Strobel et al., 2004; Congdon ym., 2008). Joustavuuteen kuuluu kyky tehdä vaihtoehtoinen vaste sen jälkeen, kun nykyinen vaste on onnistuneesti estetty. Tähänastiset havainnot inhibition ja DRD4: n välisestä yhteydestä ovat vaihtelevia. Congdon ym. (2008) havaitsi, että osallistujat, joilla on DRD4: n 7R-alleeli, osoittavat korkeamman stop-signaalin reaktioajan (SSRT) go/no-go-tehtävässä, mikä heijastaa heikompaa estävää valvontaa, kun taas Forbes et al. (2009) ei löytänyt tällaista vaikutusta Barratt Impulsiivisuusasteikkoon (bis), joka on impulsiivisuutta mittaava self-report-työkalu. Toisaalta, Colzato et al. (2010) osoitti, että nämä sekavaikutukset voivat johtua siitä, että aiemmat tutkimukset käsittelivät impulsiivisuutta monoliittisena prosessina, kun taas impulsiivisuus voidaan itse asiassa jakaa toiminnallisiin ja toimintahäiriöisiin tyyppeihin Dickmanin impulsiivisuus-inventaarion mukaan (DII; Dickman, 1990). Toimintahäiriöinen impulsiivisuus on taipumus toimia ennakoimattomasti tapauksissa, joissa tällainen toiminta on sopimatonta, kun taas toiminnallinen impulsiivisuus on samanlainen taipumus toteuttaa sopivissa tilanteissa (Colzato et al., 2010). Colzato ym. (2010) havaitsi, että DRD4 7R: llä, joka liittyy korkeampiin striataalisiin DA-tasoihin, oli korkeampi toimintahäiriöinen impulsiivisuus. Useat meta-analyyttiset tutkimukset ovat osoittaneet DRD4: n 7R-alleelin riskialttiiksi alleeliksi attention-deficit hyperactivity disorder (ADHD; Faraone et al., 2001; DiMaio et al., 2003), häiriö, jolle on ominaista korkea toimintahäiriöinen impulsiivisuus (Young et al., 2007). Lisäksi ADHD: n ja luovuuden suhdetta on tutkittu useissa tutkimuksissa (Shaw and Brown, 1990; Healey ja Rucklidge, 2006), joskin toistaiseksi nämä tutkimukset ovat tuottaneet epäselviä tuloksia. Vaikka jotkut ovat havainneet ADHD: n ja sen oireiden korreloivan positiivisesti luovuuden kanssa (Healey and Rucklidge, 2006; White and Shah, 2011), toiset ovat raportoineet päinvastaisesta suuntauksesta (Funk et al., 1993; Healey ja Rucklidge, 2005, 2008). Mielenkiintoista on, että ADHD liittyy käyttäytymiseen, joka heikentää toimeenpanovalvontaa, mukaan lukien attentional set-shifting (Boonstra et al., 2005), ja joustavuus (Barkley et al., 1997; kersantti ym., 2002), jotka molemmat ovat tärkeitä kognitiiviselle joustavuudelle, luovuuden keskeiselle ulottuvuudelle (Dietrich, 2004; Durstewitz and Seamans, 2008). Nämä havainnot osoittavat, että yksilöt, joilla on DRD4: n 7R-alleeli, saattavat osoittaa vähäisempää luovuutta ja vähentynyttä joustavuutta erityisesti DT-tehtävissä. Näin ollen DRD4 exon III VNTR on biologisesti uskottava ehdokas edistää yksilöllisiä eroja luovuutta.
yhdessä näyttää siltä, että yhtäältä 7R-alleeli, joka on riskialleeli johtotehtävien heikkenemiselle, joustavuudelle ja joukon siirtämiselle, saattaa liittyä vähäiseen luovuuteen. Toisaalta 7R-alleelin ja korkeamman uutuudenhakuisuuden välinen yhteys osoittaa, että yksilöt, joilla on 7R-alleeli, saattavat itse asiassa osoittaa suurempaa joustavuutta ja luovuutta.
nykyisen tutkimuksen tarkoituksena oli tutkia DRD4 7R: n monimutkaista roolia ihmisen luovuuden osittaisessa muokkaamisessa. Jotta voitaisiin paremmin kuvata tätä suhdetta, arvioimme kahdenlaisia DT-testejä (figural ja verbal). Kunkin tehtävän pisteytys jaettiin DT: n kolmen pääulottuvuuden, eli luovan sujuvuuden, joustavuuden ja omaperäisyyden mukaan, ja jokainen ulottuvuus analysoitiin erikseen testatakseen hypoteesia, jonka mukaan DRD4 vaikuttaa eri tavoin jokaiseen luovuuden ulottuvuuteen.
menetelmä
osallistujat
otokseen osallistui 185 opiskelijaa Haifan yliopistosta Israelista (112 naista). Kaikki osallistujat olivat valkoihoisia (itse ilmoittautuneita) ja heidän keski-ikänsä oli 24, 5 (SD ± 2, 1). Osallistujia värvättiin yliopistolla julkaistujen ilmoitusten kautta. Kaikki osallistujat olivat palkattuja vapaaehtoisia.
luovuuden arviointi
luovuutta arvioitiin kahdella DT-testillä, jotka olivat osa-asteikkoa luovan ajattelun Torrance-testin (Torrance, 1974) figuraalisesta osakokeesta, ja vaihtoehtoisessa käytössä tehtävä (AUT; Guilford et al., 1978). Molemmat testit sisälsivät DT: n kolme keskeistä ulottuvuutta eli joustavuuden, omaperäisyyden ja sujuvuuden.
Torrance Test of Creative Thinking (Circles Sub-Scale)
osallistujille annettiin sivu, jolle piirrettiin 30 identtistä ympyrää. Heitä pyydettiin piirtämään mahdollisimman monta erilaista merkityksellistä kohdetta 10 minuutin aikarajan sisällä, jolloin jokaisen piirroksen on sisällettävä vähintään yksi ympyrä. Pisteytykseen kuuluivat sujuvuus (tuotettujen vastausten määrä), joustavuus (luokkien lukumäärä) ja omaperäisyys, jotka laskettiin alkuperäisten vastausten pisteytyksen mukaan, kuten yksityiskohtaisesti Torrance Tests of Creative Thinking scoring guide-julkaisussa (Torrance, 1974).
Alternative Uses Task (AUT)
osallistujille annettiin viiden yleisen objektin luettelo ja heitä pyydettiin luettelemaan mahdollisimman monta vaihtoehtoista käyttötarkoitusta kullekin objektille 10 minuutin kuluessa. Yleisin Arkikäyttö oli merkitty suluissa. Esineet olivat: kenkä (yleinen käyttö: kulumista jalka); nappi (yleinen käyttö: sulkeminen asioita); lyijykynä (yleinen käyttö: piirustus tai kirjoitus); rengas (yleinen käyttö: auton pyörä); ja juomalasi (yleinen käyttö: sisältää nestettä). Ainoastaan vastaukset, joissa ei toistettu annettuja yleisiä käyttötarkoituksia, laskettiin ja otettiin huomioon. Torrancen testin tapaan pisteytykseen kuuluivat sujuvuus, joustavuus ja omaperäisyys. Koska alkuperäisten vasteiden pisteyttämiseen AUT: ssa ei ole ohjeita, alkuperäiset vasteet määriteltiin tilastollisesti harvinaisiksi vasteiksi laboratoriossamme suoritetun esitestin mukaan.
Pretest
asianmukaisen vastetiheyden kriteerin luomiseksi 100 terveen osallistujan ryhmä, joka ei osallistunut tähän tutkimukseen, suoritti AUT-kokeen. Jokaiselle kohteelle kerättiin lista kaikista mahdollisista käyttötarkoituksista kaikilta osallistujilta. Tämän luettelon perusteella laskettiin tilastollinen harvinaisuusaste, jotta voitiin arvioida kunkin vastauksen ja myöhemmin kunkin osallistujan omaperäisyyspisteet. Vastauksille annettiin nolla pistettä, jos 5% tai useampi osallistujista listasi tietyn käyttötarkoituksen, yksi pistemäärä, Jos 2% – 4,99% osallistujista listasi sen, ja kaksi pistettä, jos alle 1,99% listasi käytön. Kullekin osallistujalle laskettiin keskimääräinen alkuperäisyyspistemäärä näiden tilastollisten harvinaisuuspisteiden perusteella.
DNA: n uuttaminen ja genotyypin määritys
DNA uutettiin 20 ml: sta suuvesinäytteitä Master Pure kit-menetelmällä (Epicentre, Madison, WI). DRD4 48-bp VNTR: lle oli ominaista PCR – monistusmenetelmä seuraavilla alukkeilla: F5′ – CTT CTT ACC CCT CAT GCT GCT GCT GCT CTGG – 3′ ja R5′ – ACC ACCACC GGC AGG ACC CTC ATG GCC TTG CGC TC-3. PCR-reaktiot suoritettiin käyttäen 5 µl Master Mixiä (Thermo scientific), 2 µl alukkeita (0,5 µM), 0,6 µl Mg/Cl2 (2,5 mM), 0.4 µl DMSO 5% ja 1 µl vettä yhteensä 9 µl kokonaistilavuuteen ja lisäksi 1 µl genomista DNA: ta lisättiin seokseen. Kaikki PCR-reaktiot suoritettiin Biometra T1-Termosyklaattorilla (Biometra, Güttingem, Saksa). PCR-reaktiotila oli seuraava: esilämmitysvaihe 94,0°C: ssa 5 minuutin ajan, 34 denaturointisykliä 94,0°C: ssa 30 sekunnin ajan, uudelleenjuonti 55°C: ssa 30 sekunnin ajan ja laajennus 72°C: ssa 90 sekunnin ajan. reaktio eteni 72°C: ssa 5 minuutin ajan. Tämän jälkeen reaktioseos elektroforoitiin 3-prosenttisella agaroosigeelillä (AMRESCO) etidiumbromidilla genotyyppien seulomiseksi.
genotyypin frekvenssin jakauma oli Hardy-Weinbergin tasapainon mukainen (chi-neliö = 0, 01, p-arvo = 0, 92).
tilastollinen analyysi
genotyyppi luokiteltiin DRD4: n 7R-alleelin esiintymisen tai puuttumisen perusteella (7 vs. no7), kuten kirjallisuudessa on aiemmin raportoitu (Ding ym., 2002). Ryhmien erot luovuuspisteissä analysoitiin tilastollisesti ANOVAs-menetelmällä, DRD4 7R-alleelin läsnäolo (7) tai puuttuminen (No7) itsenäisenä muuttujana. Tutkiaksemme DT: n eri näkökohtia (sujuvuus, joustavuus, omaperäisyys) suoritimme varianssin (MANOVA) monimuuttujaanalyysin erikseen kullekin komponentille.
tulokset
varmistaaksemme, että nämä kaksi ryhmää (7, no7) eivät poikenneet merkittävästi toisistaan iän ja koulutuksen suhteen, teimme riippumattomia t-testejä näiden kahden ryhmän vertailemiseksi (KS.taulukko 1 keskiarvot ja keskihajonnat). Tämä analyysi paljasti, että nämä kaksi ryhmää eivät eroa toisistaan iän tai koulutusvuosien suhteen. Ei–parametriset testit (Mann-Whitney) osoittivat merkittäviä eroja sukupuolen frekvenssijakaumassa (Z = 2, 11, p < 0, 05).
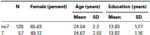
Taulukko 1. Ikä, sukupuoli ja koulutus tiedot DRD4 7R genotyyppi, SD: keskihajonta.
kahdessa tehtävässä käytettyjen luovuusmittareiden keskiarvot on esitetty taulukossa 2.

Taulukko 2. Kahden käytetyn luovuustehtävän keskiarvot ja keskivirhe (SE) : AUT ja Torrance.
Sujuvuuskomponentti
monimuuttujainen ANOVA osoitti DRD4 7R-genotyypin yleisen vaikutuksen sujuvuuskomponentille, F(2,182) = 3, 25, p<0, 05. Kun otetaan huomioon kokonaistestin merkitys, tutkittiin yhtenäisiä päävaikutuksia. Merkittävät univariaattiset päävaikutukset DRD4-genotyypille saatiin aut-sujuvuudella F(1 183) = 4, 95, p < 0.05(7R-alleeli, jolla oli pienempi sujuvuus no7-ryhmään verrattuna, Kuva 1a), mutta ei saavuttanut Torrancen sujuvuuden kannalta merkittävää tasoa F (1,183) = 2,95, p = 0,087.
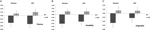
kuva 1. Luovuuspisteet AUT: n ja Torrancen tehtävistä erotetaan sujuvuuteen (a), joustavuuteen (B) ja omaperäisyyteen (C). Keinot muutettiin Z-pisteiksi.
Joustavuuskomponentti
Joustavuusmittareiden monimuuttuja-Anova vaikutti yleisesti DRD4 7R-genotyyppiin F(2,182) = 4.38, p < 0, 05. Merkittäviä univariaattisia päävaikutuksia DRD4-genotyypille saatiin aut-joustavuudelle F (1 183) = 4, 57, p < 0, 05 ja Torrance-joustavuudelle F(1 183) = 6, 95, p < 0, 01. Kuten kuviosta 1b käy ilmi, molemmissa tapauksissa 7R-alleeliryhmän yksilöillä oli pienempi joustavuus kuin No7-ryhmän yksilöillä (Taulukko 2).
alkuperäisyyskomponentilla
Alkuperäisyysmittareiden monimuuttuja-ANOVALLA ei ollut merkitseviä vaikutuksia, F(2,182) = 2,06, p = 0.13, vaikka tulokset eivät osoittavat pienempi omaperäisyys 7R alleeli, DRD4 (katso kuva 1c, Taulukko 2 lisätietoja).
sukupuolierot
Keskustelu
DT-tehtävät edellyttävät osallistujilta useita ratkaisuja tiettyyn ongelmaan. Tässä käytetyissä tehtävissä osallistujien piti miettiä monia vaihtoehtoisia käyttötapoja jokapäiväisille esineille (AUT: ssa) ja piirtää useita piirroksia, joissa on tietty muoto (ympyrä) (Torrance-tehtävässä). Tässä esitetyt tulokset osoittavat, että DRD4 7R-alleelin omaavilla yksilöillä on pienempi luovuus DT: llä mitattuna. Ottaen huomioon, että 7R-alleeli on yhdistetty toimintahäiriöiseen impulsiivisuuteen ja huonoon inhibitioon (Congdon et al., 2008; Dreber et al., 2009), se, että yksilöt, joilla on 7R-alleeli, osoittavat vähemmän luovuutta, voidaan selittää kyvyttömyydellään tukahduttaa tai estää ilmeisiä (yhteisiä) vasteita, kuten alhaisemmat omaperäisyyspisteet osoittavat. Pienempi joustavuus-ja sujuvuuspistemäärä voi viitata siihen, että kun yhteinen vaste on annettu, on vaikeampi siirtyä uuteen luokkaan (kuten alhaisen joustavuuden pisteet osoittavat), mikä johtaa Vähemmän vastauksia (kuten alhainen sujuvuuspistemäärä osoittaa). Lisätodisteita luovuuden ja impulsiivisuuden suhteesta voidaan nähdä 7R-alleelin yhteydestä ADHD: hen (Li et al., 2006), sekä lisääntynyt impulsiivisuus ADHD-potilailla (Langley et al., 2004). Mielenkiintoista on, että luovuus on yhdistetty da-järjestelmään skitsofrenian tapauksessa (Eysenck, 1993). Vaikka useat tutkimukset osoittavat, että skitsofreniaa ja skitsotyyppistä persoonallisuutta sairastavilla potilailla on suuri luovuus (O ’ Reilly et al., 2001; Folley and Park, 2005), viimeaikaiset skitsofrenian ja luovuuden mallit tukevat ylösalaisin käännettyä U-muotoista yhdistystä, jossa luovuus voi olla korkeampi matalan keskivaikean skitsotyyppisen piirteen kanssa, mutta vähenee skitsofrenian psykopatologian vakavuuden kasvaessa (Tsakanikos and Claridge, 2005; Stoneham and Coughtrey, 2009; Nelson and Rawlings, 2010). Itse asiassa tuoreessa tutkimuksessa Jaracz et al. (2012) raportoi, että skitsofreniapotilailla on vähentynyt luovuus verrattuna kontrolleihin. Lisäksi kirjoittajat kertoivat, että heikentynyt luovuus liittyi heikkoon kognitiiviseen joustavuuteen ja impulsiivisuuteen, mikä näkyi heidän huonoina tuloksinaan Wisconsinin Korttilajittelutestissä (WCST). Nämä tulokset ovat linjassa aiemman tutkimuksen kanssa, joka osoittaa, että kroonista skitsofreniaa sairastavilla potilailla on vähemmän luovuutta, joka luovan sujuvuuden tapauksessa välittyi pisteillä toimeenpanevan valvonnan tehtävistä ja omaperäisyyden tapauksessa ei (Abraham et al., 2007). Yhdessä, vaikka lievä toiminnallinen impulsiivisuus voi parantaa luovuutta potilailla, joilla on skitsotyyppisiä piirteitä, erityisesti sellaisten ideoiden tuottamisessa, jotka poikkeavat annetuista esimerkeistä (Abraham and Windmann, 2008), vaikeassa skitsofreniassa havaittu häiriintynyt impulsiivisuus voi heikentää luovia kykyjä.
nykyisen tutkimuksen tulokset tukevat luovuuden ja Da-geneettisten polymorfismien varianttien välistä suhdetta (Reuter et al., 2006; Runco ym., 2011). Lisäksi näytteestämme löydetyt tulokset osoittavat, että DRD4-7R-alleelin tärkein vaikutus luovuuteen voi johtua sen vaikutuksesta joustavuuteen. Useat tutkimukset ovat yhdistäneet kognitiivisen joustavuuden DA-polkuihin (Ashby and Isen, 1999; Dreisbach and Guschke, 2004; Müller et al., 2007). On oletettu, että D2: n kaltaisten reseptorien (eli D2: n, D3: n ja D4: n) aktivaatio vähentää Gabaergistä inhibitiota ja helpottaa useiden representaatioiden aktivoitumista, mikä lisää joustavuutta, mikä ilmenee setinsiirtokykyinä (Müller et al., 2007; Seamans and Robbins, 2010). Lisäksi D2: n kaltaiset reseptorit on yhdistetty uuden tiedon joustavaan integrointiin (Durstewitz and Seamans, 2008). Tämän mukaisesti Reuter et al. (2006) löysi dopamiinireseptorin D2: n ja luovuuden välisen yhteyden yhdistelmäindeksillä mitattuna, vaikka sitä, oliko joustavuudella merkittävä rooli näissä tuloksissa, ei voida määrittää. Lisäksi Runco et al. (2011) raportoi eroista luovissa sujuvuudessa useiden dopamiinigeenien kantajien, muun muassa DRD4: n, välillä. Heidän raportistaan ei kuitenkaan käy ilmi, kummalla ryhmällä on enemmän luovia kykyjä. On tärkeää huomata, että toisin kuin tässä esitetyt tulokset, Runco et al. (2011) ei löytänyt yhteyttä DT: n tehtävien ja DRD4: n mittaamien joustavuustoimenpiteiden välillä. Tämä epäjohdonmukaisuus voi selittyä käytettyjen populaatioiden ja ryhmäkokojen eroilla tai otokseen valitun populaation alleelifrekvenssien eroilla. Lisäksi on näyttöä siitä, että dopamiinin ja DT: n välinen yhteys ei välttämättä ole lineaarinen (Akbari Chermahini and Hommel, 2010, 2012), mikä osoittaa, että useiden DA-geenien välinen vuorovaikutus voi vaikuttaa kunkin geneettisen vaihtelun yksilöllisiin tuloksiin DT: n yhteydessä. Lisäksi useat tutkimukset ovat viitanneet siihen, että DA polut voivat olla sukupuolisensitiivisiä (Tammimäki ja Männistö, 2011; Wang et al., 2012). Zhong ym. (2010) esimerkiksi havaitsi, että sukupuoli moduloi DRD4 ekson3 polymorfismin ja oikeudenmukaisuuden tunteen välistä yhteyttä. Nykyisessä tutkimuksessa Emme löytäneet vuorovaikutusvaikutusta DRD4-7R: n ja sukupuolen välillä, mikä osoittaa, että sukupuolella ei ole moduloivaa vaikutusta DRD4: n ja luovuuden yhdistymiseen otoksessamme. Lisäksi tässä esitetyt tulokset osoittavat, että sukupuolten väliset erot luovuudessa ja erityisesti DT: ssä puuttuvat (KS.Baer and Kaufman, 2008).
johtopäätöksenä tässä tutkimuksessa esitetään, että DRD4: n ja luovuuden välillä löytyneeseen yhteyteen vaikuttaa lähinnä joustavuus. Nykyisen tutkimuksen tulokset ovat linjassa aiempien tulosten kanssa, jotka viittaavat DA-järjestelmän osallistumiseen luovuuteen ja lisäävät kertyvää tietoa, joka viittaa geneettisiin vaikutuksiin luovuudessa.
tutkimuksessa on useita rajoituksia, jotka on otettava huomioon. Ensinnäkin nykyisessä tutkimuksessa tarkasteltiin vain da—järjestelmän yhden geneettisen vaihtelun—DRD4: n-ja DT: n välistä yhteyttä. Striataalisessa DA-järjestelmässä on kuitenkin muita tärkeitä polymorfismeja, jotka voivat liittyä DT: hen ja voivat vaikuttaa sekä itsenäisesti että geenien ja geenien välisissä vuorovaikutuksissa. Yksi tällainen polymorfismi on dopamiinikuljettaja (DAT1), jonka on osoitettu liittyvän paluun estoon (Colzato et al., 2010) sekä impulsiivisuuden toimenpiteillä (Gizer ja Waldman, 2012). Lisäksi, kuten edellä mainittiin, toinen mahdollisuus, jota ei tutkittu tässä on mahdollinen geenin ja geenin vuorovaikutus vaikutuksia DT. Akbari Chermahini and Hommel (2010) raportoi epälineaarisen yhteyden striataalisen dopamiinin ja DT: n merkkiaineen välillä, mikä osoittaa, että suhde voi olla monimutkaisempi siten, että yksi geenin polymorfismi voi vaikuttaa DT: hen toisen geenin polymorfismin välityksellä. Siksi tulevissa tutkimuksissa olisi harkittava näiden kahden polymorfismin vuorovaikutuksen tutkimista joustavuuden ja DT: n yhteydessä.
Eturistiriitalausunto
kirjoittajat toteavat, että tutkimus tehtiin ilman kaupallisia tai taloudellisia suhteita, joita voitaisiin pitää mahdollisena eturistiriitana.
Akbari Chermahini, S., and Hommel, B. (2010). (B) yhteys luovuuden ja dopamiinin välillä: spontaanit silmänräpäyksnopeudet ennustavat ja dissosioivat toisistaan poikkeavaa ja konvergenttia ajattelua. Kognitio 115, 458-465. doi: 10.1016 / J.kognitio.2010.03.007
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Cools, R. (2008). Dopamiinin rooli käyttäytymisen motivoivassa ja kognitiivisessa kontrollissa. Neurotieteilijä 14, 381-395. doi: 10.1177/1073858408317009
Pubmed Abstract | Pubmed kokoteksti | CrossRef kokoteksti
Frank, M. J. (2005). Dynaaminen dopamiinimodulaatio tyvitumakkeessa: neurokomputationaalinen selvitys kognitiivisista vajeista lääkkeellisissä ja lääkkeettömissä Parkinsonismeissa. J. Cogn. Neurotieteilijä. 17, 51–72. doi: 10.1162/0898929052880093
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Healey, D., and Rucklidge, J. J. (2006). Tutkimus ADHD-oireiden, luovuuden ja neuropsykologisen toiminnan suhteesta lapsilla. Lasten Neuropsykooli. 12, 421–438. doi: 10.1080/09297040600806086
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Runco, M. A., and Richards, R. (1997). Arvostettua luovuutta, arjen luovuutta ja terveyttä. Greenwich, TT: Ablex.
Torrance, E. (1974). Erot eivät ole alijäämiä. Opettaa. Coll. Rec. 75, 471–487.