- Introduzione
- Metodo
- Partecipanti
- Valutazione della creatività
- Torrance Test of Creative Thinking (Circles Sub-Scale)
- Alternate Uses Task (AUT)
- Pretest
- Estrazione e genotipizzazione del DNA
- Analisi statistica
- Risultati
- Il componente di fluidità
- La componente di flessibilità
- La componente di originalità
- Differenze di genere
- Discussione
- Dichiarazione sul conflitto di interessi
Introduzione
cognizione Creativa gioca un ruolo importante nelle arti, nella invenzione e innovazione, così come nella vita di tutti i giorni (Runco e Richards, 1997). Sebbene la creatività sia stata considerata una capacità umana unica, i comportamenti spontaneamente creativi (ad es., creando nuovi strumenti tra primati e uccelli) hanno dimostrato di verificarsi anche nei non umani (Byrne e Bates, 2007), attestando ulteriormente le profonde radici evolutive e biologiche della creatività. La creatività è stata definita come la capacità di produrre risposte che sono sia nuove (cioè originali, rare e inaspettate) che adatte (cioè adattive e utili in base ai vincoli del compito) (Sternberg e Lubart, 1999). Uno degli approcci psicometrici alla misurazione della creatività coinvolge compiti di pensiero divergente (DT) in cui ai partecipanti viene chiesto di rispondere a un dato problema con più soluzioni (Dietrich e Kanso, 2010). I test di DT generalmente misurano vari aspetti della creatività, tra cui fluidità creativa, flessibilità e originalità (Torrance, 1974). Pertanto, sebbene non sia sinonimo di creatività, i compiti DT forniscono misurazioni strutturate e oggettive della creatività (Sternberg e Lubart, 1999; Jung et al., 2009). In particolare, i punteggi sui compiti DT hanno dimostrato di essere correlati positivamente con misure ecologicamente valide di realizzazione creativa (Carson et al., 2005) e con creatività autovalutata (Furnham e Bachtiar, 2008).
Studi precedenti hanno evidenziato il coinvolgimento del sistema dopaminergico (DA) nella creatività (Heilman et al., 2003; Flaherty, 2005; Takeuchi et al., 2010). Takeuchi et al. (2010) ha trovato differenze individuali nella creatività, misurate da DT, per essere positivamente correlate con la materia grigia nelle regioni del sistema DA, tra cui la corteccia prefrontale dorsolaterale, i gangli basali bilaterali, la substantia nigra e l’area ventrale tegmentale. Inoltre, diversi studi genetici hanno dimostrato una relazione tra DT e neurotrasmissione della dopamina (Reuter et al., 2006; De Manzano et al., 2010; Runco et al., 2011). Ad esempio, Reuter et al. (2006) ha trovato la creatività, misurata dai compiti DT che coinvolgono sia la creatività figurale che verbale, per essere significativamente associata ai polimorfismi del gene del recettore D2 della dopamina (DRD2). Inoltre, diversi studi hanno trovato che la dopamina corticale è coinvolta nella flessibilità cognitiva (Frank, 2005; Cools, 2008; Garcia-Garcia et al., 2010), uno dei componenti principali di DT.
Un gene candidato DA interessante per la creatività è il gene del recettore della dopamina D4 (DRD4). Il recettore DRD4 è uno dei cinque recettori della dopamina e svolge un ruolo importante nella mediazione della segnalazione sinaptica della dopamina. Il gene è caratterizzato da un numero variabile di 48 coppie di basi di ripetizioni tandem (48-bp VNTR) situato nella regione codificante del terzo esone. DRD4 48-bp VNTR polimorfismo varia da 2 a 11 ripetizioni tra individui (Asghari et al., 1995; Cravchik e Goldman, 2000), dove la ripetizione 4 (4R) è la ripetizione più comune nelle popolazioni caucasiche e la ripetizione 7 (7R) è la seconda variante più comune (Chang et al., 1996; Ding et al., 2002). È interessante notare che il 7R è stato precedentemente associato a comportamenti creativi della vita reale, come il tratto di personalità in cerca di novità (Ebstein et al., 1996). In effetti, la ricerca della novità-la tendenza verso l’attività esplorativa—è considerata una delle caratteristiche delle persone creative (Chavez—Eakle et al., 2006; Drago et al., 2009). Sebbene le prove di un’associazione tra DRD4 7R e la ricerca di novità siano incoerenti, come evidenziato da diversi rapporti di meta-analisi (Kluger et al., 2002; Munafò et al., 2008), molti studi hanno trovato un’associazione significativa tra la ricerca di novità e l’allele 7R del DRD4 (Ebstein et al., 1996; Benjamin et al., 2000; Keltikangas-Järvinen et al., 2002; Becker et al., 2005). Roussos et al. (2009) hanno recentemente suggerito che le differenze nella misurazione della ricerca di novità utilizzando le scale di self-report possono spiegare i risultati incoerenti tra il polimorfismo 7R e la ricerca di novità. In linea con ciò, è stato ripetutamente riscontrato che il sistema DA svolge un ruolo importante nel tratto della personalità della ricerca della novità (Flaherty, 2005; Schweizer, 2006), attestando ulteriormente il potenziale ruolo del DRD4 nella creatività.
Tuttavia, in contrasto con il ruolo di DRD4 48-bp VNTR 7R nella ricerca di novità, prove recenti indicano che 7R è in realtà associato a una ridotta flessibilità cognitiva, uno degli aspetti principali della creatività (Strobel et al., 2004; Congdon et al., 2008). La flessibilità implica la capacità di effettuare una risposta alternativa dopo aver inibito con successo una risposta corrente. I risultati finora sull’associazione tra inibizione e DRD4 sono misti. Congdon et al. (2008) ha scoperto che i partecipanti con l’allele 7R del DRD4 mostrano un tempo di reazione del segnale di arresto più elevato (SSRT) su un’attività go/no-go, riflettendo un controllo inibitorio più povero, mentre Forbes et al. (2009) non è riuscito a trovare un tale effetto sulla Barratt Impulsiveness Scale (BIS), uno strumento di auto-report per misurare l’impulsività. D’altra parte, Colzato et al. (2010) ha dimostrato che questi effetti misti possono essere dovuti al fatto che studi precedenti hanno affrontato l’impulsività come un processo monolitico, mentre l’impulsività può essere effettivamente divisa in tipi funzionali e disfunzionali secondo l’inventario di impulsività di Dickman (DII; Dickman, 1990). L’impulsività disfunzionale è la tendenza ad agire senza accortezza nei casi in cui tale azione è inappropriata, mentre l’impulsività funzionale è una tendenza simile implementata in situazioni appropriate (Colzato et al., 2010). Colzato et al. (2010) ha scoperto che gli individui con DRD4 7R, che è associato a livelli più elevati di DA striatale, hanno mostrato una maggiore impulsività disfunzionale. Diversi studi meta-analitici hanno indicato l’allele 7R del DRD4 come un allele di rischio per il disturbo da deficit di attenzione e iperattività (ADHD; Faraone et al., 2001; DiMaio et al., 2003), un disturbo caratterizzato da elevata impulsività disfunzionale (Young et al., 2007). Inoltre, la relazione tra ADHD e creatività è stata studiata in diversi studi (Shaw e Brown, 1990; Healey e Rucklidge, 2006), anche se finora questi studi hanno dato risultati poco chiari. Mentre alcuni hanno trovato l’ADHD e la sua sintomatologia positivamente correlata alla creatività (Healey e Rucklidge, 2006; White e Shah, 2011), altri hanno riportato una tendenza opposta (Funk et al., 1993; Healey e Rucklidge, 2005, 2008). È interessante notare che l’ADHD è comportamentalmente associato a menomazioni nel controllo esecutivo, incluso il set-shifting attentivo (Boonstra et al., 2005), e flessibilità (Barkley et al., 1997; Sergeant et al., 2002), entrambi importanti per la flessibilità cognitiva, una dimensione centrale della creatività (Dietrich, 2004; Durstewitz e Seamans, 2008). Questi risultati indicano che gli individui con l’allele 7R del DRD4 possono mostrare livelli più bassi di creatività e diminuita flessibilità sui compiti DT in particolare. Quindi, il DRD4 exon III VNTR è un candidato biologicamente plausibile per contribuire alle differenze individuali nella creatività.
Collettivamente, sembra che da un lato l’allele 7R, come allele di rischio per funzioni esecutive compromesse, flessibilità e set shifting, possa essere associato a bassa creatività. D’altra parte, l’associazione tra l’allele 7R e una maggiore ricerca di novità indica che gli individui con l’allele 7R possono effettivamente mostrare maggiore flessibilità e creatività.
L’attuale studio è stato progettato per esplorare il complesso ruolo del DRD4 7R nel modellare parzialmente la creatività umana. Per meglio caratterizzare questa relazione, abbiamo valutato due tipi di test DT (figurale e verbale). Il punteggio di ogni attività è stato diviso in base alle tre dimensioni principali di DT, vale a dire fluidità creativa, flessibilità e originalità, e ogni dimensione è stata analizzata separatamente per testare l’ipotesi che DRD4 influenzi in modo differenziato ogni dimensione della creatività.
Metodo
Partecipanti
Il campione comprendeva 185 studenti dell’Università di Haifa in Israele (112 donne). Tutti i partecipanti erano caucasici (auto-segnalati), con un’età media di 24,5 (DS ± 2,1). I partecipanti sono stati reclutati attraverso annunci pubblicati presso l’università. Tutti i partecipanti sono stati pagati volontari.
Valutazione della creatività
La creatività è stata valutata utilizzando due test DT, una sotto-scala dal sotto-test figurale del Torrance test of creative thinking (Torrance, 1974), e il compito di usi alternativi (AUT; Guilford et al., 1978). Entrambi i test includevano le tre dimensioni principali di DT, ovvero flessibilità, originalità e fluidità.
Torrance Test of Creative Thinking (Circles Sub-Scale)
Ai partecipanti è stata data una pagina su cui sono stati disegnati 30 cerchi identici. È stato chiesto loro di disegnare quanti più oggetti significativi possibili entro un limite di tempo di 10 min, in cui ogni disegno deve includere almeno un cerchio. Il punteggio includeva fluidità (numero di risposte prodotte), flessibilità (numero di categorie) e originalità, calcolata in base al punteggio delle risposte originali, come dettagliato nei test di Torrance of Creative Thinking scoring guide (Torrance, 1974).
Alternate Uses Task (AUT)
Ai partecipanti è stato dato un elenco di cinque oggetti comuni e chiesto di elencare quanti più usi alternativi possibili per ogni oggetto, entro un limite di tempo di 10 min. L’uso quotidiano più comune è stato indicato tra parentesi. Gli oggetti erano: scarpa (uso comune: indossare a piedi); pulsante (uso comune: chiusura cose); matita (uso comune: disegno o scrittura); pneumatico (uso comune: ruota auto); e bicchiere (uso comune: contiene liquido). Sono state contate e incluse solo le risposte che non hanno ribadito gli usi comuni indicati. Come nel test Torrance, il punteggio includeva fluidità, flessibilità e originalità. Poiché non ci sono linee guida per il punteggio delle risposte originali nell’AUT, le risposte originali sono state definite come risposte statisticamente poco frequenti secondo un pre-test condotto nel nostro laboratorio.
Pretest
Allo scopo di creare un valido criterio di frequenza di risposta, un gruppo di partecipanti sani 100 che non hanno preso parte a questo studio ha completato l’AUT. Per ogni oggetto, è stato raccolto un elenco di tutti i possibili usi da tutti i partecipanti. Sulla base di questo elenco è stata calcolata una misura statistica di infrequenza al fine di valutare il punteggio di originalità per ciascuna risposta e, successivamente, per ciascun partecipante. Alle risposte è stato assegnato un punteggio pari a zero se il 5% o più dei partecipanti ha elencato un determinato uso, un punteggio di uno se tra il 2% e il 4,99% dei partecipanti lo ha elencato e un punteggio di due se meno dell ‘ 1,99% ha elencato l’uso. Un punteggio medio di originalità è stato calcolato per ogni partecipante in base a questi punteggi statistici di scarsa frequenza.
Estrazione e genotipizzazione del DNA
Il DNA è stato estratto da 20 ml di campioni di collutorio utilizzando il Master Pure kit (Epicentre, Madison, WI). Il DRD4 48-bp VNTR era caratterizzato da una procedura di amplificazione PCR con i seguenti primer: F5′ – CTT CCT ACC CTG CCC GCT CAT GCT GCTGCT CTA CTGG – 3′ e R5′ – ACC ACC GGC AGG ACC CTC ATG GCC TTG CGC TC – 3. Le reazioni di PCR sono state condotte usando 5 µl Master Mix (Thermo scientific), 2 µl primer (0,5 µM), 0,6 µl Mg/Cl2 (2,5 mM), 0.4 µl DMSO 5% e 1 µl di acqua per un totale di 9 µl di volume totale e un ulteriore 1 µl di DNA genomico è stato aggiunto alla miscela. Tutte le reazioni di PCR sono state effettuate su un termociclatore Biometra T1 (Biometra, Güttingem, Germania). La condizione di reazione della PCR era la seguente: fase di preriscaldamento a 94,0°C per 5 min, 34 cicli di denaturazione a 94,0°C per 30 s, reannealing a 55°C per 30 s ed estensione a 72°C per 90 s. La reazione ha continuato a rimanere a 72°C per 5 min. La miscela di reazione è stata quindi elettroforizzata su un gel di agarosio al 3% (AMRESCO) con bromuro di etidio per esaminare i genotipi.
La distribuzione della frequenza del genotipo era secondo l’equilibrio di Hardy-Weinberg (chi-quadrato = 0,01, valore p = 0,92).
Analisi statistica
Il genotipo è stato classificato in base alla presenza o all’assenza dell’allele 7R del DRD4 (7 vs. no7), come in linea con precedenti rapporti in letteratura (Ding et al., 2002). Le differenze di gruppo sui punteggi di creatività sono state analizzate statisticamente usando ANOVA, con la presenza (7) o l’assenza (no7) dell’allele DRD4 7R come variabile indipendente. Per esaminare i diversi aspetti della DT (fluidità, flessibilità, originalità), abbiamo condotto un’analisi multivariata della varianza (MANOVA) separatamente per ciascun componente.
Risultati
Per confermare che i due gruppi (7, no7) non erano significativamente diversi in termini di età e istruzione, abbiamo condotto test t indipendenti per confrontare i due gruppi (vedi Tabella 1 per i mezzi e le deviazioni standard). Questa analisi ha rivelato che i due gruppi non differivano in termini di età o anni di istruzione. I test non parametrici (Mann–Whitney) hanno rivelato differenze significative nella distribuzione di frequenza per genere (Z = 2.11, p < 0.05).
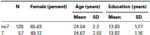
Tabella 1. Età, sesso e istruzione dati per il genotipo DRD4 7R, SD: deviazione standard.
I punteggi medi delle misure di creatività utilizzate nei due compiti sono riassunti nella tabella 2.

Tabella 2. Mezzi e errore standard (SE) dei due compiti di creatività utilizzati: AUT e Torrance.
Il componente di fluidità
Un ANOVA multivariato indicava un effetto generale del genotipo DRD4 7R per il componente di fluidità, F(2.182) = 3.25, p < 0.05. Dato il significato del test complessivo, sono stati esaminati gli effetti principali univariati. Significativi effetti principali univariati per il genotipo DRD4 sono stati ottenuti per AUT fluency F(1.183) = 4.95, p < 0.05 (allele 7R che mostra una fluidità inferiore rispetto al gruppo no7, Figura 1A), ma non ha raggiunto il livello di significatività per Torrance fluency F(1,183) = 2.95, p = 0.087.
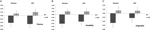
Figura 1. I punteggi della creatività sui compiti AUT e Torrance sono separati da fluidità (A), flessibilità (B) e originalità (C). I mezzi sono stati trasformati in punteggi Z.
La componente di flessibilità
ANOVA multivariata delle misure di flessibilità ha mostrato un effetto generale per DRD4 7R genotipo F(2.182) = 4.38, p < 0.05. Effetti principali univariati significativi per il genotipo DRD4 sono stati ottenuti per AUT flexibility F(1.183) = 4.57, p < 0.05 e per Torrance flexibility F(1.183) = 6.95, p < 0.01. Come mostrato nella Figura 1B, in entrambi i casi gli individui del gruppo allele 7R hanno mostrato una flessibilità inferiore rispetto agli individui del gruppo no7 (Tabella 2).
La componente di originalità
ANOVA multivariata delle misure di originalità non ha mostrato alcun effetto significativo, F(2,182) = 2.06, p = 0.13, sebbene i punteggi indicassero una minore originalità per l’allele 7R del DRD4 (vedi Figura 1C, Tabella 2 per i dettagli).
Differenze di genere
Discussione
Le attività DT richiedono ai partecipanti di fornire più soluzioni a un dato problema. Nelle attività utilizzate qui, ai partecipanti è stato richiesto di pensare a molti usi alternativi possibili per gli oggetti di uso quotidiano (nell’AUT) e di disegnare più disegni che incorporano una forma specifica (cerchio) (nell’attività Torrance). I risultati presentati qui dimostrano che gli individui con l’allele DRD4 7R mostrano una creatività inferiore misurata da DT. Considerando che l’allele 7R è stato associato ad impulsività disfunzionale e scarsa inibizione (Congdon et al., 2008; Dreber et al., 2009), il fatto che gli individui con l’allele 7R esibiscano una creatività inferiore può essere spiegato dalla loro incapacità di sopprimere o inibire risposte ovvie (comuni), come indicato da punteggi di originalità inferiori. I punteggi di flessibilità e fluidità inferiori possono indicare che una volta data una risposta comune, diventa più difficile passare a una nuova categoria (come indicato dai punteggi di flessibilità bassa), portando a meno risposte (come indicato dal punteggio di fluidità bassa). Ulteriori prove per la relazione tra creatività e impulsività possono essere viste nell’associazione dell’allele 7R con l’ADHD (Li et al., 2006), così come con una maggiore impulsività nei pazienti con ADHD (Langley et al., 2004). È interessante notare che la creatività è stata collegata al sistema DA nel caso della schizofrenia (Eysenck, 1993). Sebbene diversi studi dimostrino che i pazienti con schizofrenia e personalità schizotipica esibiscono un’alta creatività (O’Reilly et al., 2001; Folley e Park, 2005), i recenti modelli di schizofrenia e creatività supportano un’associazione a forma di U invertita in cui la creatività può essere più alta con tratto schizotipico basso-moderato, ma diminuisce con l’aumentare della gravità della psicopatologia nella schizofrenia (Tsakanikos e Claridge, 2005; Stoneham e Coughtrey, 2009; Nelson e Rawlings, 2010). Infatti, in un recente studio Jaracz et al. (2012) ha riferito che i pazienti con schizofrenia mostrano una ridotta creatività rispetto ai controlli. Inoltre, gli autori hanno riferito che la creatività compromessa era associata a una bassa flessibilità cognitiva e impulsività, riflessa dai loro punteggi bassi sul Wisconsin Card Sorting Test (WCST). Questi risultati sono in linea con precedenti ricerche che dimostrano che i pazienti con schizofrenia cronica mostrano una minore creatività, che nel caso della fluidità creativa era mediata da punteggi sui compiti di controllo esecutivo e nel caso dell’originalità non lo era (Abraham et al., 2007). Collettivamente, mentre l’impulsività funzionale lieve può migliorare la creatività tra i pazienti con tratti schizotipici, specialmente nella generazione di idee che differiscono dagli esempi forniti (Abraham e Windmann, 2008), l’impulsività disfunzionale osservata nella schizofrenia grave può smorzare le capacità creative.
I risultati dell’attuale studio supportano la relazione tra creatività e varianti dei polimorfismi genetici DA (Reuter et al., 2006; Runco et al., 2011). Inoltre, i risultati trovati nel nostro campione indicano che l’effetto principale dell’allele DRD4-7R sulla creatività può essere dovuto al suo impatto sulla flessibilità. Diversi studi hanno collegato la flessibilità cognitiva con i percorsi DA (Ashby e Isen, 1999; Dreisbach e Goschke, 2004; Müller et al., 2007). È stato postulato che l’attivazione dei recettori simili a D2 (cioè D2, D3 e D4) diminuisca l’inibizione gabaergica e faciliti l’attivazione di rappresentazioni multiple, aumentando così la flessibilità manifestata dalle capacità di set-shifting (Müller et al., 2007; Seamans e Robbins, 2010). Inoltre, i recettori simili a D2 sono stati collegati all’integrazione flessibile di nuove informazioni (Durstewitz e Seamans, 2008). In linea con questo, Reuter et al. (2006) ha trovato un’associazione tra il recettore della dopamina D2 e la creatività misurata da un indice composito, sebbene non sia possibile determinare se la flessibilità abbia svolto un ruolo importante in questi risultati. Inoltre, Runco et al. (2011) ha riportato differenze nella fluidità creativa tra i portatori di diversi geni della dopamina, tra cui il DRD4. Tuttavia, non è chiaro dal loro rapporto quale gruppo esibisce maggiori capacità creative. È importante notare che, a differenza dei risultati presentati qui, Runco et al. (2011) non ha trovato un’associazione tra le misure di flessibilità misurate dai compiti DT e DRD4. Questa incoerenza può essere spiegata da differenze nelle popolazioni utilizzate e nelle dimensioni del gruppo o da differenze nelle frequenze degli alleli all’interno della popolazione campionata. Inoltre, ci sono prove che suggeriscono che l’associazione tra dopamina e DT potrebbe non essere lineare (Akbari Chermahini e Hommel, 2010, 2012), indicando che un’interazione tra diversi geni DA può influenzare i singoli risultati di ciascuna variabilità genetica nel contesto di DT. Inoltre, diversi studi hanno sottolineato il fatto che i percorsi DA possono essere sensibili al genere (Tammimäki e Männistö, 2011; Wang et al., 2012). Zhong et al. (2010), ad esempio, ha scoperto che il genere modulava l’associazione tra polimorfismo dell’esone DRD4 e senso di equità. Nello studio attuale non abbiamo trovato un effetto di interazione tra il DRD4-7R e il genere, indicando che il genere non ha un effetto modulante sull’associazione tra DRD4 e creatività nel nostro campione. Inoltre, i risultati presentati qui indicano una mancanza di differenze di genere nella creatività, e in particolare nel DT (per la revisione, vedere Baer e Kaufman, 2008).
In conclusione, nell’attuale studio suggeriamo che l’associazione trovata tra DRD4 e creatività sia principalmente influenzata dalla flessibilità. I risultati dell’attuale studio sono in linea con i risultati precedenti che indicano il coinvolgimento del sistema DA nella creatività e aggiungono al corpo di conoscenze che si accumula suggerendo influenze genetiche nella creatività.
Ci sono diverse limitazioni dello studio che dovrebbero essere riconosciute. Innanzitutto, l’attuale studio ha esaminato solo l’associazione tra una variabilità genetica nel sistema DA—il DRD4—e il DT. Tuttavia, ci sono altri polimorfismi importanti nel sistema DA striatale che potrebbero essere associati a DT e possono contribuire sia in modo autonomo che nelle interazioni gene-gene. Uno di questi polimorfismi è il trasportatore della dopamina (DAT1), che ha dimostrato di essere associato all’inibizione del ritorno (Colzato et al., 2010) così come con misure di impulsività (Gizer e Waldman, 2012). Inoltre, come accennato in precedenza, un’altra possibilità che non è stata esplorata qui sono i possibili effetti di interazione gene-gene sulla DT. Akbari Chermahini e Hommel (2010) hanno riportato un’associazione non lineare tra un marker di dopamina striatale e DT, indicando che la relazione potrebbe essere più complessa, in modo tale che un polimorfismo di un gene possa influenzare il DT attraverso la mediazione di un altro polimorfismo genico. Pertanto, gli studi futuri dovrebbero considerare l’esame dell’interazione tra questi due polimorfismi nel contesto della flessibilità e del DT.
Dichiarazione sul conflitto di interessi
Gli autori dichiarano che la ricerca è stata condotta in assenza di relazioni commerciali o finanziarie che potrebbero essere interpretate come un potenziale conflitto di interessi.
Akbari Chermahini, S., e Hommel, B. (2010). Il (b) legame tra creatività e dopamina: i tassi di lampeggio spontaneo degli occhi predicono e dissociano il pensiero divergente e convergente. Cognizione 115, 458-465. doi: 10.1016 / j. cognizione.2010.03.007
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Cools, R. (2008). Ruolo della dopamina nel controllo motivazionale e cognitivo del comportamento. Neuroscienziato 14, 381-395. doi: 10.1177/1073858408317009
Pubmed Abstract | Pubmed Full Text / CrossRef Full Text
Frank, MJ (2005). Modulazione dinamica della dopamina nei gangli della base: un resoconto neurocomputazionale dei deficit cognitivi nel parkinsonismo medicato e non medicato. J. Cogn. Neurosci. 17, 51–72. doi: 10.1162/0898929052880093
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Healey, D., and Rucklidge, JJ (2006). Un’indagine sulla relazione tra sintomatologia ADHD, creatività e funzionamento neuropsicologico nei bambini. Neuropsicolo infantile. 12, 421–438. doi: 10.1080 / 09297040600806086
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Runco, MA, and Richards, R. (1997). Creatività eminente, creatività quotidiana e salute. A. S., CT: Ablex.
Torrance, E. (1974). Le differenze non sono deficit. Insegnare. Coll. Rec. 75, 471–487.