はじめに
創造的認知は、芸術、発明および革新、ならびに日常生活において重要な役割を果たす(Runco and Richards、1997)。 創造性はユニークな人間の能力と考えられてきましたが、自発的に創造的な行動(例えば、 霊長類や鳥の間で新しいツールを作成する)は、非ヒトにも発生することが示されている(Byrne and Bates、2007)、創造性の深い進化と生物学的ルーツをさらに証明している。 創造性は、新規(すなわち、オリジナル、希少および予期しない)および適切(すなわち、タスクの制約に従って適応性があり有用)の両方である応答を生成す 創造性を測定するための心理測定的アプローチの1つには、参加者が複数の解決策で特定の問題に対応するように求められる発散的思考(DT)タスクが DTのテストは、一般的に創造性の様々な側面を測定します,創造的な流暢さを含みます,柔軟性と独創性(Torrance,1974). したがって、創造性と同義ではないが、DTタスクは、創造性の構造化された客観的な測定を提供する(Sternberg and Lubart,1 9 9 9;Jung e t a l., 2009). 特に、DTタスクのスコアは、創造的達成の生態学的に有効な尺度と正の相関があることが示されている(Carson et al. 2005)だけでなく、自己評価された創造性(FurnhamとBachtiar、2008)。
これまでの研究では、ドーパミン作動性(DA)システムが創造性に関与していることが指摘されている(Heilman et al.,2003;Flaherty,2005;Takeuchi et al., 2010). 竹内他 (2010)は、DTによって測定された創造性の個人差が、背外側前頭前野、両側大脳基底核、黒質、および被蓋腹側領域を含むDA系領域における灰白質と正の相関を さらに、いくつかの遺伝学的研究は、DTとドーパミン神経伝達との間の関係を示している(Reuter e t a l. ら、2 0 0 6;De Manzano e t a l. ら,2 0 1 0;Runco e t a l., 2011). 例えば、Reuter e t a l. (2006)は、図と言葉の両方の創造性を含むDTタスクによって測定される創造性が、ドーパミンD2受容体遺伝子(DRD2)の多型と有意に関連していることを さらに、いくつかの研究では、皮質ドーパミンが認知の柔軟性に関与していることが見出されている(Frank、2005;Cools、2008;Garcia-Garcia et al.,2010),DTの主要なコンポーネントの一つ.
創造性のための興味深いDA候補遺伝子の一つは、ドーパミンD4受容体遺伝子(DRD4)です。 DRD4受容体は五つのドーパミン受容体の一つであり、シナプスドーパミンシグナル伝達を仲介する上で重要な役割を果たしている。 この遺伝子は、第三のエクソンのコード領域に位置するタンデムリピート(48bp VNTR)の48塩基対可変数によって特徴付けられる。 DRD4 4 8−bp VNTR多型は、個体間で2〜1 1回の反復を変化させる(Asghari e t a l. ら、1 9 9 5;CravchikおよびGoldman、2 0 0 0)、ここで、4反復(4R)は、白人集団において最も一般的な反復であり、7反復(7R)は、2番目に最も一般的な変異体である(Chang e t a l.,1 9 9 5;CravchikおよびGoldman、2 0 0 0)。 ら、1 9 9 6;Ding e t a l., 2002). 興味深いことに、7Rは、以前に、新規性を求める性格特性などの現実の創造的行動に関連していた(Ebstein et al., 1996). 確かに、新規性を求めること-探索的活動への傾向—は、創造的な人々の特徴の一つであると考えられている(Chavez—Eakle et al. ら,2 0 0 6;Drago e t a l., 2009). DRD4 7Rと新規性探索との間の関連についての証拠は矛盾しているが、いくつかのメタ分析報告によって証明されるように(Kluger e t a l. ら,2 0 0 2;Munafu e t a l. ら、2 0 0 8)、多くの研究は、新規性探索とDRD4の7R対立遺伝子との間に有意な関連性を見出した(Ebstein e t a l.,2 0 0 8)。 ら、1 9 9 6;Benjamin e t a l. ら、2 0 0 0;Keltikangas−Jarvinenら、2 0 0 1)。 ら,2 0 0 2;Becker e t a l., 2005). Roussos et al. (2009)は最近、自己報告尺度を用いた新規性探索の測定の違いが、7R多型と新規性探索の間の矛盾した結果を説明する可能性があることを示唆している。 これに沿って、DAシステムが新規性を求める性格特性において主要な役割を果たしていることが繰り返し判明しており(Flaherty、2005;Schweizer、2006)、創造性におけるDRD4の潜在的な役割をさらに証明しています。それにもかかわらず、新規性探索におけるDRD4 48-bp VNTR7Rの役割とは対照的に、最近の証拠は、7Rが実際には創造性の主要な側面の1つである認知柔軟性の障害と関連していることを示している(Strobel et al. ら、2 0 0 4;Congdon e t a l., 2008). 柔軟性には、現在の応答を正常に阻害した後に代替応答を行う能力が含まれる。 阻害とDRD4との関連に関するこれまでの知見は混合されている。 Congdon et al. (2 0 0 8)は、DRD4の7R対立遺伝子を有する参加者が、進行中/進行中での停止信号反応時間(SSRT)が高く、抑制制御が貧弱であることを反映していることを見出し、Forbes e t a l. (2009)は、衝動性を測定するための自己報告ツールであるBarratt Impulsiveness Scale(BIS)にこのような効果を見つけることができなかった。 一方、Colzato et al. (2010)は、これらの混合効果は、以前の研究が衝動性をモノリシックプロセスとして扱っていたという事実に起因する可能性があることを実証したが、衝動性は実際にはDickmanの衝動性インベントリ(DII;Dickman,1990)に従って機能的タイプと機能不全タイプに分けられる可能性がある。 機能不全衝動性は、そのような行動が不適切な場合に先見の明なく行動する傾向であり、機能的衝動性は適切な状況で実施される同様の傾向である(Colzato et al., 2010). Colzato et al. (2010)は、より高いレベルの線条体DAに関連するDRD4 7Rを有する個体が、より高い機能不全衝動性を示すことを見出した。 いくつかのメタ分析的研究は、注意欠陥多動性障害のリスク対立遺伝子としてDRD4の7R対立遺伝子を指摘している(ADHD;Faraone e t a l. ら、2 0 0 1;Dimaio e t a l. ら、2 0 0 3)、高い機能不全衝動性を特徴とする障害(Young e t a l., 2007). さらに、ADHDと創造性との関係は、いくつかの研究で調査されている(Shaw and Brown、1990; これまでのところ、これらの研究は不明な結果をもたらしているが、Healey and Rucklidge,2006)。 ADHDとその症状が創造性と正の相関があることを発見した人もいますが(Healey and Rucklidge、2006;White and Shah、2011)、反対の傾向を報告している人もいます(Funk et al. ら、1 9 9 3;Healey and Rucklidge,2 0 0 5,2 0 0 8)。 興味深いことに、ADHDは、注意セットシフトを含む、行動的に実行制御の障害と関連している(Boonstra et al. ら、2 0 0 5)、およびflexibity(Barkley e t a l. ら、1 9 9 7;Sergaeら、1 9 9 8;S. 両方とも認知の柔軟性にとって重要であり、創造性の中心的な次元である(Dietrich、2004;Durstewitz and Seamans、2008)。 これらの知見は、DRD4の7R対立遺伝子を有する個人が、特にDTタスクに対するより低いレベルの創造性および減少した柔軟性を示す可能性があるこ したがって、DRD4エクソンIII VNTRは、創造性の個人差に貢献するための生物学的にもっともらしい候補です。
集合的に、一方では、7R対立遺伝子は、障害された実行機能、柔軟性およびセットシフトのリスク対立遺伝子として、低い創造性と関連している可能性があるようである。 一方、7R対立遺伝子とより高い新規性探索との間の関連は、7R対立遺伝子を有する個人が実際により大きな柔軟性および創造性を示すことを示
現在の研究は、人間の創造性を部分的に形成する際のDRD4 7Rの複雑な役割を探求するために設計されました。 この関係をよりよく特徴付けるために、我々は2つのタイプのDTテスト(図と口頭)を評価した。 各タスクのスコアリングは、DTの三つの主要な次元、すなわち創造的な流暢さ、柔軟性と独創性に応じて分割され、各次元は、DRD4は、差別的に創造性の各次元に影響を与えるという仮説をテストするために別々に分析されました。
メソッド
参加者
サンプルは、イスラエルのハイファ大学から185人の学生(112人の女性)で構成されていました。 すべての参加者は白人(自己報告)であり、平均年齢は24.5(SD±2.1)であった。 参加者は、大学で掲示された発表を通じて募集されました。 すべての参加者は有料のボランティアでした。
創造性の評価
創造性は、創造的思考のトーランステスト(Torrance、1974)のfiguralサブテストからのサブスケール、および代替使用タスク(AUT;Guilford et al., 1978). 両方のテストには、DTの3つのコア次元、すなわち柔軟性、独創性、流暢さが含まれていました。
創造的思考のトーランステスト(サークルサブスケール)
参加者には、30個の同一の円が描かれたページが与えられました。 彼らは、10分の制限時間内にできるだけ多くの異なる意味のあるオブジェクトを描画するように求められました。 スコアリングには、Torrance Tests of Creative Thinking scoring guide(Torrance、1974)に詳述されているように、元の回答のスコアリングに従って計算された流暢さ(回答の数)、柔軟性(カテゴリの数)、独創性が含まれていました。
Alternate Uses Task(AUT)
参加者には五つの共通オブジェクトのリストが与えられ、10分の制限時間内に各オブジェクトについてできるだけ多くの代替 最も一般的な日常使用は括弧内に示されていました。 オブジェクトは、靴(一般的な使用:徒歩で着用)、ボタン(一般的な使用:閉じるもの)、鉛筆(一般的な使用:描画または書き込み)、タイヤ(一般的な使用:車の車輪)、お 与えられた一般的な用途を繰り返し表明しなかった回答のみがカウントされ、含まれました。 トーランス試験と同様に、得点には流暢さ、柔軟性、独創性が含まれていました。 AUTでは元の応答のスコアリングのガイドラインがないため、元の応答は、私たちの研究室で実施された事前テストに従って統計的にまれな応答と定義されていました。
Pretest
応答頻度の有効な基準を作成する目的で、この研究に参加しなかった100人の健康な参加者のグループがAUTを完了しました。 各オブジェクトについて、すべての可能な使用のリストがすべての参加者から収集されました。 統計的低頻度尺度は、各回答の独創性スコアを評価するために、このリストに基づいて計算され、その後、各参加者のために。 回答には、参加者の5%以上が特定の用途を記載している場合はゼロのスコアが割り当てられ、参加者の2%から4.99%の間に記載されている場合は1のスコア、1.99%未満が用途を記載している場合は2のスコアが割り当てられました。 これらの統計的低頻度スコアに従って、各参加者について平均独創性スコアを計算した。Dna抽出および遺伝子型決定(genotyping)</h4><p>DNAを、master Pure kit(Epicentre,Madison,W I)を使用して、2 0mlのうがい薬サンプルから抽出した。</p><h4>DNA抽出およびgenotyping(genotyping)</h4><P>DNAを、 DRD4 4 8−bp VNTRを、以下のプライマー:F5’−CTT CCT ACC CTG CCC GCT CAT GCT GCTGCT CTA CTGG−3’およびR5’−ACC ACCACC GGC AGG ACC CTC ATG GCC TTG CGC TC−3を用いたPCR増幅手順によって特徴付けた。 PCR反応は、5μ lのMaster Mix(Thermo scientific)、2μ lのプライマー(0.5μ M)、0.6μ lのMg/Cl2(2.5m M)、0を用いて行った。4μ lのDMSO5%および1μ lの水を合計して合計体積9μ lにし、さらに1μ lのゲノムDNAを混合物に添加した。 PCR反応は全て、Biometra T1Thermocycler(Biometra,Guttingem,Germany)で行った。 PCR反応条件は以下の通りであった:94.0℃で5分間予熱ステップ、94.0℃で30秒間34サイクルの変性、55℃で30秒間再アニール、および72℃で90秒間延長。 次いで、反応混合物を臭化エチジウムを含む3%アガロースゲル(AMRESCO)上で電気泳動し、遺伝子型をスクリーニングした。遺伝子型頻度の分布は、ハーディ-ワインバーグ平衡(カイ二乗=0.01、p値=0.92)に従っていた。
遺伝子型頻度の分布は、ハーディ-ワインバーグ平衡に従っていた(カイ二乗=0.01、p値=0.92)。
統計分析
遺伝子型は、文献における以前の報告に沿ったように、DRD4の7R対立遺伝子の有無(7対no7)に従って分類された(Ding et al., 2002). 創造性スコアのグループの違いは、独立変数としてDRD4 7R対立遺伝子の存在(7)または不在(no7)で、ANOVAsを使用して統計的に分析しました。 DTの異なる側面(流暢性,柔軟性,独創性)を調べるために,各成分について多変量分散分析(MANOVA)を別々に行った。
結果
二つのグループ(7、no7)は、年齢と教育の面で有意に異ならなかったことを確認するために、我々は二つのグループを比較するために独立したt検定を実施した(平均と標準偏差については表1を参照)。 この分析は、二つのグループは、年齢や教育の年の面で異ならなかったことを明らかにしました。 ノンパラメトリック検定(マン-ホイットニー)は、性別の頻度分布に有意差を明らかにした(Z=2.11、p<0.05)。テーブル1
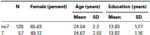
テーブル1。 DRD4 7R遺伝子型の年齢、性別および教育データ、SD:標準偏差。
2つのタスクで使用される創造性尺度の平均スコアを表2にまとめました。テーブル2

テーブル2。 使用される2つの創造性タスクの平均と標準誤差(SE): AUTとトーランス
流暢成分
多変量ANOVAは、流暢成分F(2,182)=3.25,p<0.05に対するDRD4 7R遺伝子型の一般的な効果を示しました。 全体的なテストの重要性を考慮して、一変量の主な効果を調べた。 DRD4遺伝子型の有意な一変量主効果は、AUT流暢F(1,183)=4.95,p<0のために得られた。05(7R対立遺伝子はno7群と比較して低い流暢性を示す、図1A)が、トーランス流暢性F(1,183)=2.95、p=0.087の有意水準に達しなかった。
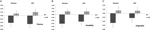
図1。 AUTとトーランスのタスクの創造性スコアは、流暢さ(A)、柔軟性(B)、独創性(C)に分かれています。 Zスコアに変換されました。
柔軟性成分
柔軟性尺度の多変量ANOVAは、DRD4 7R遺伝子型F(2,182)=4に対して一般的な効果を示した。38,p<0.05. DRD4遺伝子型の有意な一変量主効果は、AUT柔軟性F(1,183)=4.57、p<0.05とトーランス柔軟性F(1,183)=6.95、p<0.01のために得られた。 図1Bに示すように、両方の場合において、7R対立遺伝子群の個体は、no7群の個体と比較して低い柔軟性を示した(表2)。
独創性成分
独創性尺度の多変量ANOVAは有意な効果を示さなかった、F(2,182)=2.06、p=0。図13に示すように、スコアはDRD4の7R対立遺伝子の独創性が低いことを示していたが(詳細については図1C、表2を参照)。
性差
議論
DTタスクでは、参加者は特定の問題に対して複数の解決策を提供する必要があります。 ここで使用されたタスクでは、参加者は、(AUTで)日常のアイテムのための多くの代替可能な用途を考えると、(トーランスタスクで)特定の形状(円)を組み込んだ複数の図面を描くために必要とされました。 ここに提示された結果は、DRD4 7r対立遺伝子を有する個体が、DTによって測定されるように、より低い創造性を示すことを実証する。 7r対立遺伝子が、機能不全の衝動性および不十分な阻害と関連していることを考慮すると(Congdon e t a l.,2 0 0 2)、7r対立遺伝子は、機能不全の衝動性および不十分 ら、2 0 0 8;Dreber e t a l. ら、2009)、7R対立遺伝子を有する個体がより低い創造性を示すという事実は、より低い独創性スコアによって示されるように、明らかな(一般的な)応答を抑 柔軟性と流暢性のスコアが低いと、共通の回答が与えられると、新しいカテゴリに移行することが難しくなり(柔軟性の低いスコアで示される)、回答が少なくなる(流暢性の低いスコアで示される)ことを示すことがある。 創造性と衝動性との間の関係についてのさらなる証拠は、7R対立遺伝子とADHDとの関連において見ることができる(Li e t a l. ら、2 0 0 6)、ならびにADHD患者における衝動性の増加を伴う(Langley e t a l., 2004). 興味深いことに、創造性は統合失調症の場合のDAシステムにリンクされています(Eysenck、1993)。 いくつかの研究は、統合失調症および統合失調症の人格を有する患者が高い創造性を示すことを示すが(O’Reilly et al. 統合失調症および創造性の最近のモデルは、創造性が低中等度の分裂型形質で高くなるが、統合失調症における精神病理学の重症度が増加すると減少する反転U字型の関連性を支持する(Tsakanikos and Claridge、2005;Stoneham and Coughtrey、2009;Nelson and Rawlings、2010)。 実際、最近の研究では、Jaracz et al. (2012)は、統合失調症の患者が対照と比較して創造性の低下を示すことを報告した。 さらに、Wisconsin Card Sorting Test(WCST)のスコアが低いことが反映されているため、創造性の障害は認知の柔軟性と衝動性の低下と関連していることが報告されている。 これらの結果は、慢性統合失調症の患者がより低い創造性を示すことを示す以前の研究に沿ったものであり、創造的な流暢さの場合には執行制御タス, 2007). 集合的に、軽度の機能的衝動性は、特に与えられた例とは異なるアイデアを生成する際に、分裂型形質を有する患者の創造性を改善する可能性がある(Abraham and Windmann、2008)が、重度の統合失調症で観察される機能不全衝動性は創造的能力を弱める可能性がある。
現在の研究の結果は、創造性とDA遺伝子多型の変異体との関係を支持している(Reuter et al. ら,2 0 0 6;Runco e t a l., 2011). さらに、我々のサンプルで見つかった結果は、創造性に対するDRD4-7r対立遺伝子の主な効果は、柔軟性への影響によるものである可能性があることを示 いくつかの研究は、認知の柔軟性をD A経路と関連づけている(Ashby and Isen,1 9 9 9;Dreisbach and Goschke,2 0 0 4;Munller e t a l., 2007). D2様受容体(すなわち、D2、D3およびD4)の活性化は、Gaba作動性阻害を減少させ、複数の表現の活性化を容易にし、したがってセットシフト能力によ ることができます。 さらに、D2様受容体は、新しい情報の柔軟な統合にリンクされている(Durstewitz and Seamans、2008)。 これに沿って、Reuter e t a l. (2006)は、複合指数によって測定されたドーパミン受容体D2と創造性との間の関連を見出したが、柔軟性がこれらの結果に大きな役割を果たしたかどうかを決定することはできない。 さらに、Runco e t a l. (2011)は、いくつかのドーパミン遺伝子のキャリア間で創造的な流暢さの違いを報告しました,それらの間でDRD4. しかし、どのグループがより創造的な能力を発揮するかは、彼らの報告書から明らかではない。 ここで提示された結果とは対照的に、Runco et al. (2011)は、DTタスクとDRD4によって測定された柔軟性の尺度との間に関連性を見出さなかった。 この不一致は、使用された集団およびグループサイズの違い、またはサンプリングされた集団内の対立遺伝子頻度の違いによって説明され得る。 さらに、ドーパミンとDTの間の関連付けが線形のものではないかもしれないことを示唆する証拠があり(Akbari Chermahini and Hommel、2010、2012)、いくつかのDA遺伝子間の相互作用がDTの文脈における各遺伝的変動の個々の結果に影響を与える可能性があることを示している。 さらに、いくつかの研究は、DA経路が性別に敏感である可能性があるという事実を指摘している(Tammimäki and Männistö、2011;Wang et al。, 2012). Zhongら。 (2010)は、例えば、性別がDRD4エクソン3多型と公平性の感覚との間の関連を調節することを見出した。 現在の研究では、DRD4-7Rと性別の間に相互作用効果は見られず、性別はサンプルのDRD4と創造性の関連に変調効果を持たないことを示しています。 さらに、ここで提示された結果は、創造性、特にDTにおける性差の欠如を示している(レビューについては、Baer and Kaufman、2008を参照)。結論として、現在の研究では、DRD4と創造性の間に見られる関連は、主に柔軟性によって影響されることを示唆している。 現在の研究の結果は、創造性におけるDAシステムの関与を指摘する以前の結果と一致しており、創造性における遺伝的影響を示唆する知識の蓄積
認められるべき研究にはいくつかの制限があります。 まず、現在の研究では、DAシステムにおける1つの遺伝的変動性(DRD4)とDTとの関連のみを検討しました。 しかし、線条体DAシステムには、DTに関連する可能性があり、独立した方法だけでなく、遺伝子-遺伝子相互作用の両方に寄与する可能性のある他の重要な多型がある。 そのような多型の1つは、ドーパミン輸送体(DAT1)であり、これは、戻りの阻害に関連することが示されている(Colzato e t a l. と同様に、衝動性の尺度(Gizer and Waldman、2012)。 さらに、上述したように、ここでは検討されなかった別の可能性は、DTに対する遺伝子-遺伝子相互作用効果の可能性である。 Akbari ChermahiniとHommel(2010)は、線条体ドーパミンのマーカーとDTとの間の非線形関連を報告し、遺伝子の1つの多型が別の遺伝子多型の仲介によってDTに影響を与える可能性があるように、関係がより複雑である可能性があることを示している。 したがって、将来の研究では、柔軟性とDTの文脈でこれら二つの多型間の相互作用を調べることを検討すべきである。
利益相反に関する声明
著者らは、この研究は、利益相反の可能性と解釈される可能性のある商業的または財政的関係がない場合に行われたと宣言している。
Akbari Chermahini,S.,And Hommel,B.(2010). 創造性とドーパミンとの間の(b)リンク:自発的な目の瞬き率は、発散的および収束的思考を予測し、解離する。 115,458-465 土井:10.1016/j.認知.2010.03.007
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Cools,R.(2008). 行動の動機づけおよび認知制御におけるドーパミンの役割。 神経科学者14、381-395。 doi:10.1177/1073858408317009
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Frank,M.J.(2005). 大脳基底核における動的ドーパミン変調:薬用および非投薬パーキンソニズムにおける認知障害の神経計算アカウント。 J.Cogn. ニューロシ… 17, 51–72. doi:10.1162/0898929052880093
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Healey,D.,And Rucklidge,J.J.(2006). 子供のADHDのsymptomatology、創造性およびneuropsychological機能間の関係への調査。 子供の神経精神。 12, 421–438. doi:10.1080/09297040600806086
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Runco,M.A.,And Richards,R.(1997). 卓越した創造性、毎日の創造性、そして健康。 グリニッジ、CT:Ablex。
トーランス、E.(1974)。 違いは赤字ではありません。 教えて コッ… Rec. 75, 471–487.
div