A Microbial Biorealm page on the genus Lactobacillus delbrueckii
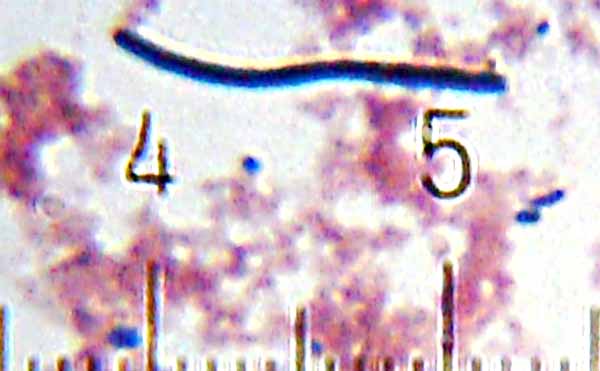
Numbered ticks are 11 µM apart.
Gram-stained.
Photograph by Bob Blaylock.
Classification
Higher order taxa
Bacteria (Domain); Firmicutes (Phylum); Bacilli (Class); Lactobacillales (Order); Lactobacillaceae(家族)
種
NCBI:分類
Lactobacillus delbrueckii
subspecies: bulgaricus, lactis, delbrueckii, and indicus
説明と意義
Lactobacillus us delbrueckiiは、棒状のグラム陽性、非運動性細菌です。 種に共通するのは、嫌気性条件下で糖基質を乳酸生成物に発酵させる能力である。 そのため、L.delbrueckiiは、l.delbrueckii subspを除いて、ヨーグルト、牛乳、チーズなどの乳製品に一般的に含まれています。 デルブリュエッキ(3) その代謝産物とこれまでに知られているその内部遺伝学によって区別される四つの亜種があります。 最も最近の受け入れられた亜種、L.delbrueckii subsp。 indicusは、インドの乳製品(1)から単離された。 対照的に、Stamen Grigorov博士はL.delbrueckii subspを単離した。 1905年にヨーグルトのサンプルからbulgaricus。
l.delbrueckiiをホモ発酵性乳酸菌(LAB)として定義する特性は、その代謝最終生成物D-乳酸塩およびL-乳酸塩によって限定されない。 L.delbrueckii subsp. bulgaricusは、改善された乳糖耐性と免疫応答(刺激する能力を含むヒトおよび動物にプロバイオティクスの影響を有することが証明されている4、5、7)。 この情報に対する過去の議論は、後者が低酸性環境およびヒト胃腸管の胃液内で生き残る能力に疑問を呈した。 L.delbrueckii subspによって生成されるホスホ多糖。 bulgaricusは、マウス(におけるマクロファージの食作用を強化する能力を持っている4)。
ゲノム構造
Lactobacillus delbrueckii subspの環状ゲノム。 BUGARICUS ATCC11842は2006年5月に完成した。 1,864,998個のヌクレオチドで構成され、それが属する乳酸菌属の他の種と比較して異常に高いG-C含有量(49%)を有する。 2,217個の遺伝子のうち、1,562個のコードがタンパク質であり、533個のコードが擬似遺伝子である(10)。 これらのようなゲノムの特徴、ならびに挿入配列パターンは、酪農産業におけるその適応を示しており、急速な進化段階の理論を支持している(11)。
タンパク質prtBとlacオペロンをコードする1,562個の遺伝子のうち、L.delbrueckiiのホモ発酵特性にとって重要です。 Lacオペロン内には、ラクトースの取り込みおよび分解をコードするlacS、lacZ、およびlacR遺伝子がある。(3)lacsの遺伝子は膜を通して乳糖を運ぶ機能に責任がある乳糖のpermeaseのためにコードします。 ラクトース代謝に必要な重要な酵素B-ガラクトシダーゼはlacz遺伝子にコードされている。 LacZの下流には調節遺伝子lacRがある。
細胞構造と代謝
グラム陽性細菌L.delbrueckiiは、グラム試験の下で紫色の染色を保持しています。 このタイプの微生物に特有のものは、厚い細胞壁と細胞膜である。 付加的な障壁として機能する外膜がないことは、バクテリオファージ攻撃に対する感受性の理由である可能性がある2)。
prtB遺伝子によってコードされるプロテアーゼは、L.delbrueckii subspの細胞壁に沿って固定されている。 bulgaricusとlactis;そしてsubspで最も可能性が高いです。 インディカス 乳製品中で成長する亜種の能力は、必須アミノ酸を暴露するカゼイン分解中の酵素活性に加えて、lacZ遺伝子の構成的または誘導性発現に起因する。 (8,4)
四つのl.delbrueckii亜種にとって重要なのは、それが代謝することができる基質の数と種類です。 前述のように、そのような特性は、そのゲノム内の酵素発現に制限される。 L.delbrueckii subsp. ブルガリクスとサブスプ。 indicusは、ラクトース、グルコース、フルクトース、およびマンノースを代謝することができます。 これらに加えて、L.delbrueckii subsp。 lactisはガラクトース、ショ糖、マルトース、trehaloseおよび他の変更された炭水化物を異化させます。(4)
生態学
発酵乳製品の住民と乳酸Lactobacillus delbrueckiiの生産者として、L.subspを除いて。 delbrueckiiは、その低酸性環境の原因です。 栄養要件は細菌の環境に適合されており、アミノ酸、ビタミン、炭水化物および不飽和脂肪酸が含まれるが、これらに限定されない(9)。 L. delbrueckiiは、嫌気性条件下で40〜44℃の最適成長温度を有する(3)。 具体的には、L.subsp. bulgaricusは、スターター乳酸菌培養物に共存するため、Streptococcus thermophilusと共生関係を持っています。
病理学
Lactobacillus delbrueckiiは非病原性である。 実際、それは食品工業で広く利用されて、ヨーグルト、ミルク、野菜およびチーズで見つけることができます。
バイオテクノロジーへの応用
これまでに知られている四つの亜種のL.delbrueckii subsp。 ブルガリクスとサブスプ。 ラクティスは、発酵乳、ヨーグルト、チーズの生産のためのスターター培養物として、酪農産業にとって最も重要です。 広く使用されているLactobacillus delbrueckii subspの発酵プロセスであれば、経済的損失は有意であろう。 ブルガリクスとサブスプ。 ラクティスは妨げられていた。 したがって、酪農産業は、バクテリオファージを検出し、安全性と貯蔵寿命(のために高品質を確保するために生産条件を調整することができなければ L.delbrueckii subspの共生関係のために。 bulgaricusとStreptococcus thermophilus後者は同時に検討されている。
現在の研究
乳酸桿菌の特定の株は、有糸分裂作用を有し、脾臓細胞増殖を助けることが示されている。 L.delbrueckii亜種bulgaricusおよびl.acidophilusの熱処理Y S株はマウスspenocytesによるIgmおよびIggの生成を直接誘導し,ATCC株とは対照的に細菌濃度に依存した。 前者は5×106および2×107乳酸菌ml–1の量で最も効果的であった。 抗体濃度をELISAおよびFisher試験で決定した。 さらに,両乳酸菌種のy s株とATCC株はリンパ球増殖を誘導した。 L.delbrueckii subsp. bulgaricusは,乳酸菌抗体の除去時に高い抗体レベルの維持から示されるポリクローナルB細胞を活性化することが示された。 (13)
ある研究では、製造の任意の段階でバクテリオファージを検出するためのマルチプレックスPCRの実用化が示唆されました。 この方法はシンプルで迅速であることが証明され、製品の最小品質要件が保証されています。 結果はLの量が低いことを示したが。 delbrueckiiファージ使用した試料中では,S.thermophilusファージの比較的高い量が見出された。 これらの結果は,スターター培養で使用されるS.thermophilusの割合の増加の結果である。 (2)
Lactobacillus delbrueckiiは、一般的に、乳製品業界でスターター培養の外に見つけることができません。 それが起源の自然環境は確かに知られていません。 最近の研究では、L.delbrueckii subspの単離と特性評価が報告されています。 伝統的なヨーグルトの準備に基づいてブルガリアの植物からの共生連鎖球菌thermophilusと一緒にbulgaricus。 ターゲット植物Cornus masと六百六十五植物サンプルは、人間の居住から離れた四つのサイトから収集されました。 L.subspの識別。 bulgaricusは表現型解析,パルスフィールドゲル電気泳動(PFGE)解析,およびPCR法により決定した。 45℃で成長した培養サンプルは、棒状であり、D-乳酸を生成し、プライマー LB1/LLB1と1065塩基対のDNA断片を生成し、タンパク質分解活性を示した。 6 6 5の植物試料から、L.delbrueckii subsp. bulgaricusとs.thermophilusは分離され,その大部分はブルガリアのBlagoevgradから来た。 (12)
1)F.,Felis,Giovanna E.,Castioni,A.,Torriani,S.,And Germond,J.”Lactobacillus delbrueckii subsp. インディカス・subsp. 【送料無料】【、インドの乳製品から分離された”。 2005. 系統的および進化的微生物学の国際ジャーナル。 第55巻 401-404頁
2)
3)J.,Lapierre,L.,Delley,M.,Mollet,B.,Felis,G.,And Dellaglio,F.”細菌Speies Lactobacillus delbrueckiiの進化: 原核生物種の概念を反映した部分的なゲノム研究”。 分子生物学と進化。 2003. 第20巻 93-104頁
4)
5)M.,Callegari,M.,Ferrari,S.,Bessi,E.,Cattibelli,D.,Soldi,S.,Morelli,L.,Feuillerat,N.,And Antoine,J.”ヒト腸におけるヨーグルト細菌の生存”. 応用環境微生物学。 2006. 第72巻 5113-5117頁
6)
7)S.,Drescher,K.,And Heller,K.”Lactobacillus delbrueckii subspの生存。 Fistulated Gottingen Minipigsの末端回腸におけるbulgaricusおよびStreptococcus thermophilus。 応用および環境微生物学。 2001. 第67巻 4137-4143頁
8)C.,D.Atlan,B.Blanc,R.Portailer,J.E.Germond,L.Lapierre,b.Mollet. 1996. “A new cell surface proteinase:Lactobacillus delbruekii subspからのprtB遺伝子の配列決定と解析。 ブルガリクス”。 細菌学のジャーナル。 1996. 第178巻 3059-3065頁
9)
10)バイオテクノロジー情報センター(NCBI)ゲノム。 Lactobacillus delbrueckii subsp. 11842
11)de Guchte、M.、Penaud、S.、Grimaldi、C.、Barbe、V.、Bryson、K.、および他。 “Lactobacillus bulgaricusの完全なゲノム配列は、広範かつ継続的な還元進化を明らかにする”。 PNAS。 2006. 第103巻 p.9274-9279
12)M.,Minkova,S.,Kimura,K.,Sasaki,T.,And Isawa,K.”Lactobacillus delbrueckii ssp. ブルガリアの植物からbulgaricusとStreptococcus thermophilus”。 FEMSの微生物学の手紙。 2007. 269巻。 160-169頁
13)
レイチェル-ラーセンのMaryruth Penetrante学生によって編集
KLBによって編集