- Inleiding
- materialen en methoden
- plantaardige materialen en Groeiconditie
- PSI-en PSII-metingen
- Elektrochromische Shiftanalyse
- foto-remmende behandelingen
- statistische analyse
- resultaten
- discussie
- de rol van CEF-stimulatie bij weinig licht
- verandering in P700-Oxidatieverhouding bij weinig licht is niet gerelateerd aan pmf
- Etrii bestuurt P700 Oxidatieverhouding bij weinig licht
- conclusie
- Auteursbijdragen
- financiering
- belangenconflict verklaring
Inleiding
hoewel licht de drijvende kracht is van fotosynthese, kan groot licht een significante fotothibitie van fotosysteem II (PSII) veroorzaken voor bladeren van schaduwgevende planten (Kitao et al., 2000; Barth et al., 2001; Krause et al., 2004; Huang et al., 2015b, 2016b, c). Echter, photoinhibited PSII herstelt snel bij weinig licht (hij en Chow, 2003; Zhang en Scheller, 2004, Allakhverdiev et al., 2005), vanwege de snelle omloopsnelheid van D1-eiwit (Aro et al., 1993; Zhang and Scheller, 2004; Allakhverdiev et al., 2005). De snelheid van PSII-herstel was verminderd na remming van de synthese van ATP via PSI of PSII (Allakhverdiev et al., 2005), wat aangeeft dat de snelle reparatie van fotobeschadigde PSII-complexen een grote hoeveelheid ATP in een korte tijd nodig heeft. Een vorige studie wees uit dat na het koelen-geïnduceerde fotoinhibitie van PSII, cyclische elektronenstroom (CEF) significant werd gestimuleerd tijdens herstel bij weinig licht (Huang et al., 2010). Er werd voorgesteld dat deze stimulatie van CEF bij weinig licht de synthese van ATP voor de snelle reparatie van PSII verbeterde (Huang et al., 2010). Er is echter meer bewijs nodig om de specifieke rol te verduidelijken van CEF-stimulatie bij weinig licht na PSII-fotoinhibitie.
tijdens CEF worden elektronen van NADPH of ferredoxine rond PSI in de plastochinonpool gefietst. Deze elektronenoverdracht is gekoppeld aan proton translocatie en genereert een protongradiënt over thylakoidmembranen (ΔpH) (Shikanai and Yamamoto, 2017). Naast de ATP-synthese is een andere functie van ΔpH de downregulatie van fotosynthetisch elektronentransport door het thylakoïdlumen te verzuren (Shikanai, 2014, 2016). Deze verordening omvat twee verschillende mechanismen: een is gekoppeld aan thermische energie dissipatie en dissipeert overtollige geabsorbeerde lichtenergie als warmte van PSII antennes (Takahashi et al., 2009), en de andere is down-regulatie van CYT B6/f complexe activiteit en controleert de snelheid van elektronenoverdracht aan PSI (Suorsa et al., 2012, 2016; Tikkanen en Aro, 2014). In angiospermen met inbegrip van Arabidopsis thaliana, werken twee wegen van PSI cyclisch elektronentransport (Shikanai, 2007). De eerste CEF-route is pgr5- / PGRL1-afhankelijke route die gevoelig is voor antimycine A (Munekage et al., 2002; Sugimoto et al., 2013), en de andere is afhankelijk van het chloroplast NADH dehydrogenase-achtige (NDH) complex (Burrows et al., 1998; Shikanai et al., 1998). De bijdrage van de pgr5-/PGRL1-afhankelijke route is belangrijker in C3-installaties. Bij hoog licht induceert de activering van pgr5-/pgrl1-afhankelijke CEF de verzuring van het thylakoïdlumen en leidt dit tot de hoge niveaus van de P700-oxidatieverhouding (Suorsa et al., 2012, 2016; Kono et al., 2014; Yamori et al., 2016). Ter vergelijking, de P700 oxidatieverhouding bij weinig licht werd weinig beïnvloed door de deficiëntie in pgr5-/PGRL1-afhankelijke CEF (Munekage et al., 2002, 2004; Kono et al., 2014). Bij matige psii-fototentoonstelling ging de stimulatie van CEF bij weinig licht gepaard met een toename van het P700-oxidatieratio (Huang et al., 2010). Het is echter onduidelijk of deze verhoogde P700-oxidatieverhouding wordt veroorzaakt door de CEF-stimulatie.
naast CEF speelt elektronenoverdracht van PSII (ETRII) een belangrijke rol bij het beïnvloeden van de redoxtoestand van PSI (Tikkanen et al., 2014; Huang et al.,2016a, c; Suorsa et al., 2016). In Pgr5 planten van A. thaliana, ernstige foto-tentoonstelling van PSII zal waarschijnlijk functioneren als de ultieme controle van fotosynthetische elektronenoverdracht waardoor het behoud van P700 optimaal geoxideerd onder overtollig licht (Tikkanen et al., 2014). Onlangs, Suorsa et al. (2016) gaf aan dat pgr5-planten een hoge P700-oxidatieverhouding vertoonden onder hoog licht wanneer de ETRII werd verminderd door knockoutmutaties te combineren in psb1 -, PsbP2 -, PsbQ1 -, PsbQ2-en psbr-loci. In gekoelde bladeren van tabak, matige PSII foto-inhibitie onderdrukt elektronen stroom naar PSI en vervolgens verhoogde de P700 oxidatieverhouding tijdens verdere koeling behandelingen (Huang et al., 2016a). Bovendien leidde de chilling-geïnduceerde fotoinhibitie van PSII tot een depressie van ETRII, die gepaard ging met een toename van de P700 oxidatieverhouding bij weinig licht (Huang et al., 2010). We veronderstellen dat de P700 oxidatieverhouding bij weinig licht voornamelijk wordt gecontroleerd door elektronenstroom uit PSII.
in ons vorige rapport merkten we op dat de schaduwgevende plant Panax notoginseng selectieve foto-inhibitie vertoonde van PSII onder hoge lichtstress. Na kortdurende blootstelling aan hoge lichtstress nam ETRII bij weinig licht aanzienlijk af, maar CEF werd aanzienlijk gestimuleerd. Tegelijkertijd nam de P700-oxidatieverhouding grotendeels toe. Onze specifieke doelstellingen waren om (1) te onderzoeken of CEF-stimulatie bij weinig licht voornamelijk de synthese van ATP vergemakkelijkt; en (2) bepalen of de toename van de P700-oxidatieverhouding bij matige psii-fotoinhibitie meer verband houdt met ETRII dan met de CEF-stimulatie of met de verandering in pmf. Om deze vragen te beantwoorden, werden intacte bladeren van Panax notoginseng behandeld bij 2258 µmol fotonen m-2 s-1 gedurende 30 minuten. Voor en na behandeling met hoog licht werden de energieverdeling in PSII, de redoxtoestand van PSI en proton motive force (pmf) bij een laag licht van 54 µmol fotonen m-2 s-1 bepaald.
materialen en methoden
plantaardige materialen en Groeiconditie
in dit onderzoek werden twee jaar oude planten van een schaduwgevende plant Panax notoginseng (Burkill) F. H. Chen ex C. Chow en W. G. Huang gebruikt voor experimenten. Deze planten werden gekweekt bij een lichtconditie van 10% zonlicht (met een maximale lichtintensiteit van ongeveer 200 µmol fotonen m-2 s-1). Voor deze planten werden geen water-en voedingsstress ervaren. Voor de fotosynthetische metingen werden 9 weken oude, volledig uitgevouwen bladeren gebruikt.
PSI-en PSII-metingen
fotosysteem I en PSII-parameters werden gemeten bij 25°C door gelijktijdig te registreren met behulp van een dubbel PAM-100-meetsysteem (Heinz Walz GmbH, Effeltrich, Duitsland). De fluorescentieparameters voor chlorofyl werden als volgt berekend: Fv / Fm = (Fm-Fo)/Fm, Y(II) = (Fm’ – Fs) / Fm’ (Genty et al., 1989), NPQ = (Fm – Fm’)/Fm’. Fo en Fm zijn de minimale en maximale fluorescentie na donkere acclimatie, respectievelijk. Fs is de aan licht aangepaste steady-state fluorescentie. Fo en Fm werden bepaald na donkere acclimatisatie gedurende 30 minuten voor en na een behandeling met hoog licht. De PSI fotosynthetische parameters werden gemeten door Dual PAM-100 gebaseerd op P700 signaal (verschil van intensiteiten van 830 en 875 nm pulsgemoduleerd meetlicht dat de fotodetector bereikt) (Klughammer en Schreiber, 2008). De P700 + signalen (P) kunnen variëren tussen een minimaal (P700 volledig gereduceerd) en een maximaal niveau (P700 volledig geoxideerd). Het maximumniveau (Pm) werd bepaald met toepassing van een verzadigingspuls (600 ms en 10000 µmol-fotonen m-2 s-1) na voorbelichting met verrood licht, en Pm werd gebruikt om de PSI-activiteit te schatten. Pm ‘ werd bepaald gelijkaardig aan Pm maar met actinisch licht in plaats van ver-rood licht. De kwantumopbrengst van PSI werd berekend als Y (I) = (Pm’- P)/Pm. De oxidatieverhouding P700 in een gegeven actinisch licht werd berekend als Y ( ND) = P/Pm. De kwantumopbrengst van de niet-fotochemische energiedissipatie van PSI als gevolg van de beperking aan de acceptorzijde werd berekend als Y (NA) = (Pm-Pm’)/Pm. De steady state waarden voor Y (II), NPQ, Y(I) en Y(ND) werden gemeten na lichtacclimatatie bij weinig licht gedurende 20 minuten.
fotosynthetische elektronenstroom door PSI en PSII werden berekend als: ETRII = Y (II) ×PPFD ×0,84 ×0,5 (Krall and Edwards, 1992), ETRI = Y (I) ×PPFD×0,84 ×0,5 (Yamori et al., 2011), waarbij wordt aangenomen dat 0,5 het deel van het geabsorbeerde licht is dat PSI of PSII bereikt, en dat 0,84 de absorptie is (de fractie van het invallende licht dat door bladeren wordt geabsorbeerd). De mate van CEF-activering werd geschat als ETRI / ETRII-ratio (Yamori et al., 2011, 2015). Opgemerkt moet worden dat de 1: 1 excitatie partitionering tussen PSI en PSII kan niet echt zijn, omdat schaduwplanten de neiging om de PSII/PSI verhouding te verhogen (Lunde et al., 2003; Tikkanen et al., 2006; Grieco et al., 2012). Verder werden in deze studie PSI-en PSII-parameters gemeten met rood actinisch licht (635 nm) Van Dual-PAM 100 (Walz, Duitsland). Omdat het Rode actinische licht de opwinding van PSII boven die van PSI begunstigt, kan de energiedistributie van lhcii aan PSII en PSI door het Rode actinische licht (Tikkanen et al., 2017).
Elektrochromische Shiftanalyse
het ECS-signaal werd gemonitord als de absorptieverandering bij 515 nm met behulp van een Dual-PAM-100 (Walz, Effeltrich, Duitsland) uitgerust met een P515/535 emitter-detector module (Walz). Het ECS-signaal werd verkregen na 20 min van verlichting bij 54 µmol fotonen m-2 s-1, Daarna werd het ECS-verval gemeten door het actinische licht gedurende 30 s uit te schakelen. de analyse van ECS dark interval relaxation kinetics (DIRKECS) werd gedaan door de methode van Sacksteder et al. (2001) en Takizawa et al. (2008). De totale pmf werd geschat op basis van de totale amplitude van het snelle verval van het ECS-signaal gedurende het donkerinterval van 300 ms. De langzame ontspanning van ECS-signaal maakte het mogelijk om de bijdrage van proton gradiënt over de thylakoidmembranen (ΔpH) te herkennen. De tijdsconstante van de eerste orde ECS ontspanning (TECS) is omgekeerd evenredig met de Proton geleidbaarheid (gH+) van het thylakoid membraan door het ATP synthase (Sacksteder and Kramer, 2000; Cruz et al., 2005). Als gevolg hiervan werd gH + geschat als de inverse van de vervaltijdconstante .
foto-remmende behandelingen
na donkere acclimatie gedurende 30 minuten werden Fv / Fm en Pm gemeten in intacte bladeren. Daarna werden deze intacte bladeren aangepast aan het licht bij 59 µmol fotonen m-2 s – 1 gedurende 20 minuten en werden parameters van chlorofyl fluorescentie, P700 signaal en ECS signaal geregistreerd. Toen werd het actinische licht veranderd in 2258 µmol fotonen m-2 s-1. Na blootstelling aan dit hoge licht gedurende 30 min, werd het actinische licht onmiddellijk veranderd in 59 µmol fotonen m-2 s – 1 en de fotosynthetische parameters werden geregistreerd na lichtacclimatatie gedurende 20 min. Tenslotte werden Fv/Fm en Pm gemeten na donkere acclimatisatie gedurende 30 minuten.
statistische analyse
de resultaten werden weergegeven als gemiddelde waarden van vijf onafhankelijke experimenten. Onafhankelijke T-test werd gebruikt bij α = 0,05 significantieniveau om te bepalen of er significante verschillen bestonden tussen verschillende behandelingen. Alle statistische analyses werden uitgevoerd met SPSS 16.0.
resultaten
tijdens fotosynthetische inductie bij 59 µmol fotonen m-2 s-1 nam ETRII geleidelijk toe en bereikte een steady state na ongeveer 18 minuten (figuur 1A). Ter vergelijking: ETRI was constant tijdens deze inductiefase (figuur 1A). Na het begin van dit lage licht was ETRI veel hoger dan ETRII. Na een fotosynthetische inductie van 20 minuten was ETRI echter lager dan ETRII. Deze resultaten suggereren de activering van CEF tijdens de beginfase van de inductie, in overeenstemming met eerdere studies (Joliot en Joliot, 2002, 2005; Makino et al., 2002). Na deze fotosynthetische inductie bij weinig licht werden bladeren gedurende 30 minuten verlicht bij een hoog licht van 2258 µmol fotonen m-2 s-1. Interessant is dat ETRII geleidelijk afnam tijdens de high-light behandeling (figuur 1B). Ondertussen was de waarde van ETRI hoger dan die van ETRII (figuur 1B), wat wijst op de activering van CEF bij groot licht. Na deze high-light behandeling werd de maximale foto-oxideerbare P700 (Pm) stabiel gehouden (figuur 2A). Ter vergelijking: de maximale fluorescentieintensiteit (Fm) daalde met 40% en de maximale kwantumopbrengst van PSII (Fv/Fm) daalde van 0,80 tot 0,65 (figuren 2B,C). Deze resultaten wijzen op de selectieve foto-inhibitie van PSII in bladeren van Panax notoginseng onder hoge lichtstress.
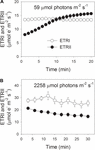
figuur 1. Fotosynthetische elektronenstroom bij weinig licht (A) en hoog licht (B). (A) donker-geacclimatiseerde bladeren werden blootgesteld aan een laag licht van 59 µmol fotonen m-2 s-1 gedurende 20 minuten. (B) na een belichting bij 59 µmol fotonen m-2 s-1 gedurende 20 min om de elektronenbak in de fotosynthese te activeren, werden de bladeren gedurende 30 min blootgesteld aan een hoog licht van 2258 µmol fotonen m-2 s-1. De waarden zijn gemiddelden ± SE (n = 5).
vóór behandeling met groot licht werden rijpe bladeren gedurende 20 minuten verlicht bij een laag licht van 59 µmol-fotonen m-2 s-1 om de fotosynthese te activeren, en vervolgens werden waarden voor ETRI, ETRII, Y(ND) en NPQ geregistreerd. Na een behandeling met hoog licht gedurende 30 minuten werden waarden voor ETRI, ETRII, Y(ND) en NPQ geregistreerd na een nieuwe acclimatiseringsfase van 20 minuten voor weinig licht. Vóór de behandeling met hoog licht waren de waarden voor ETRI en ETRII bij 59 µmol-fotonen m-2 s-1 respectievelijk 13,4 en 15,7 µmol-elektronen m-2 s-1 (figuren 3A, B). Na behandeling met hoog licht waren ETRI en ETRII bij weinig licht respectievelijk 12,8 en 11,2 µmol elektronen m-2 s-1 (figuren 3A, B). De waarde van ETRII bij 59 µmol-fotonen m-2 s-1 daalde met ongeveer 30% na foto-remmende behandeling, wat wijst op de depressie van ETRII bij matige PSII-foto-remming. Vóór de behandeling met hoog licht was de waarde van de ETRI/ETRII-verhouding bij 59 µmol-fotonen m-2 s-1 0,85 (figuur 3C). Na foto-remmende behandeling nam de ETRI / ETRII-ratio significant toe tot 1,14 (figuur 3C). Deze resultaten suggereerden de stimulatie van CEF bij weinig licht bij PSII photoinhibition (Yamori et al., 2011, 2015).
na de high-light behandeling veranderde de steady-state waarde van Y(I) bij 59 µmol fotonen m-2 s-1 niet (figuur 4A). Interessant is dat Y (NA) significant daalde van 0,36 naar 0,23(figuur 4B), en Y (ND) significant steeg van 0,1 naar 0,25 (figuur 4C). Deze resultaten wijzen op de verandering in de redoxtoestand van PSI bij weinig licht na foto-remmende behandeling. Ondertussen veranderde de waarde van NPQ iets (figuur 4D). Als gevolg hiervan had de matige psii-fotoinhibitie verschillende effecten op de P700-oxidatieverhouding en NPQ bij weinig licht.
om te verduidelijken of de toename van Y(ND) wordt veroorzaakt door een toename van pmf, werden de elektrochromische verschuivingssignalen bij 54 µmol-fotonen m-2 s-1 bepaald na lichtacclimatisatie gedurende 20 minuten vóór en na de behandeling met grootlicht. Interessant is dat de totale pmf na behandeling met groot licht met 18% aanzienlijk is afgenomen (figuur 5A), maar dat het ΔpH-niveau slechts in geringe mate is veranderd (figuur 5B). Omdat de vorming van pmf kan worden beïnvloed door de thylakoid proton geleidbaarheid, de Proton geleidbaarheid (gH+) van het thylakoid membraan bij dit lage licht werd ook onderzocht voor en na high-light behandeling. Het resultaat toonde aan dat gH+ niet veranderde met de high-light behandeling (figuur 5C), wat suggereert dat de high-light behandeling de activiteit van het chloroplast ATP synthase bij weinig licht nauwelijks beïnvloedde. Omdat de daling van 30% in ETRII gepaard ging met een daling van 18% in pmf, compenseerde de stimulatie van CEF bij weinig licht de vorming van PMF en ΔpH.

figuur 5. Waarden voor de totale protonmotive force (PMF) (A), de Proton gradiënt (ΔpH) over de thylakoïdmembranen (B), en de Proton geleidbaarheid (gH+) van het thylakoïdmembraan (C) bij weinig licht. Alle parameters werden gemeten na aanpassing bij 54 µmol fotonen m-2 s-1 gedurende 20 minuten. De waarden zijn gemiddelden ± SE (n = 5). Een sterretje geeft een significante verandering aan (P < 0,05) na behandeling met groot licht.
discussie
de rol van CEF-stimulatie bij weinig licht
Er is aangegeven dat CEF een belangrijke rol speelt bij het ondersteunen van fotosynthese en plantengroei (Yamori and Shikanai, 2016). Onder hoog licht wordt CEF nu beschouwd als essentieel voor het balanceren van het energiebudget van ATP/NADPH en voor het beschermen van PSI en PSII tegen schade (Munekage et al., 2002, 2004; Takahashi et al., 2009; Suorsa et al., 2012, 2016; Walker et al., 2014; Huang et al., 2015a, 2017b). Bij lage lichtintensiteit speelt CEF een belangrijke rol bij het optimaliseren van fotosynthetische CO2-Assimilatie waarschijnlijk via de toevoer van extra ATP (Yamori et al., 2011, 2015; Nishikawa et al., 2012; Huang et al., 2015a). De belangrijkste rol van CEF wordt flexibel gemoduleerd in reactie op de lichtintensiteit. In het bijzonder draagt CEF voornamelijk bij tot het in evenwicht brengen van het energiebudget van ATP/NADPH onder sub-verzadigde lichtsterkten, maar beschermt het fotosynthetische apparaat vooral tegen Foto-inhibitie wanneer het wordt blootgesteld aan verzadigde lichtsterkten via verzuring van het thylakoïdlumen (Huang et al., 2015a). Interessant, Huang et al. (2010) vond dat CEF beduidend bij weinig licht na het koelen-veroorzaakte photoinhibition van PSII werd gestimuleerd en veronderstelde dat deze CEF stimulatie hoofdzakelijk de synthese van ATP voor de snelle reparatie van PSII verbeterde. Er was echter meer bewijs nodig om deze hypothese te ondersteunen. In deze studie merkten we op dat de waarde van ETRI/ETRII bij weinig licht significant toenam (figuur 3C), wat wijst op de stimulatie van CEF rond PSI (Huang et al., 2011, 2012, 2017a, B; Yamori et al., 2011, 2015). Tegelijkertijd nam de amplitude van pmf significant af en nam ΔpH licht af. Uit deze resultaten bleek dat de stimulatie van CEF niet leidde tot de ΔpH-afhankelijke downregulatie van fotosynthetisch elektronentransport. Met andere woorden, deze stimulatie van CEF bij weinig licht vergemakkelijkt voornamelijk de synthese van ATP.
hoewel de protongeleiding van het chloroplastatp-synthase niet werd beïnvloed door de behandeling met hooglicht, resulteerde de kleinere grootte van de pmf in een daling van de werkelijke snelheid van ATP-synthese. De snelheid van PSII reparatie en fotosynthetische CO2 Assimilatie bij weinig licht worden voornamelijk beperkt door de snelheid van ATP synthese (Allakhverdiev et al., 2005; Yamori et al., 2011, 2015; Nishikawa et al., 2012). Omdat de ETRII significant daalde bij psii-fotoinhibitie, was de snelheid van ATP-synthese via ETRII opmerkelijk verminderd. Om de ATP/NADPH-verhouding die door primair metabolisme en de snelle reparatie van PSII wordt vereist in evenwicht te brengen, is een flexibel mechanisme nodig om extra ATP te verstrekken. Onder deze voorwaarde daalde ETRII met 30%, maar daalde de totale pmf met slechts 18%. Deze resultaten wijzen erop dat de stimulatie van CEF de vermindering van ETRII-afhankelijke vorming van pmf compenseerde, waardoor de synthese van ATP werd verbeterd. De snelle reparatie van fotobeschadigde PSII is afhankelijk van ATP synthese (Allakhverdiev et al., 2005). Versnelling van CEF rond PSI verhoogde de intracellulaire concentratie van ATP, waardoor de snelheid van PSII-reparatie in Synechocystis werd versneld (Allakhverdiev et al., 2005). Samen stellen we voor dat de stimulatie van CEF bij weinig licht een belangrijke rol speelt in de snelle reparatie van PSII activiteit via een extra ATP synthese.
verandering in P700-Oxidatieverhouding bij weinig licht is niet gerelateerd aan pmf
een interessant fenomeen is dat de P700-oxidatieverhouding bij weinig licht significant toenam bij matige psii-fotoinhibitie (Huang et al., 2010), wat ook blijkt uit de huidige studie. Onder omstandigheden waarin geabsorbeerd licht hoger is dan de vereisten voor fotosynthese, reguleert CEF-afhankelijke generatie van ΔpH de activiteit van Cyt B6/f complexe activiteit en regelt elektronen stroom van PSII naar PSI (Suorsa et al., 2012, 2016; Shikanai, 2014, 2016), het optimaliseren van de redox toestand van P700 in PSI en het minimaliseren van Ros productie in PSI tijdens fotosynthese. In Pgr5-planten van A. thaliana, wordt P700 verminderd door elektronen in het licht als gevolg van het verlies van PMF generatie. Echter, in de pgr5 mutant accumuleren flavodiiron eiwit, het niveau van pmf werd ook hersteld tot het wild-type niveau, en dus P700 werd geoxideerd als in de wild-type (Yamamoto et al., 2016). In chlorofyl B-deficiënte tarwe mutant lijnen, onvoldoende thylakoïde proton gradiënt leidt tot over-reductie van PSI acceptor zijde en dus PSI foto-inhibitie onder hoge licht of hoge temperatuur (Brestic et al., 2015, 2016). Onlangs rapporteerden sommige studies dat de verhoogde activiteit van het chloroplast ATP synthase de vorming van pmf verminderde en de over-reductie van fotosynthetische elektronentransportketen veroorzaakte, resulterend in fotoschade van PSI onder hoog licht en fluctuerend licht (Kanazawa et al., 2017; Takagi et al., 2017). Deze rapporten ondersteunen de cruciale rol van pmf bij het optimaal geoxideerd houden van P700 onder overtollige lichtenergie.
uit onze huidige resultaten blijkt dat na de behandeling met groot licht gedurende 30 minuten het totaal gevormd pmf bij dit geringe licht aanzienlijk daalde en de vorming van ΔpH lichtjes veranderde (figuren 5A,B). Ondertussen steeg de P700-oxidatieverhouding aanzienlijk van 0,1 (vóór behandeling) tot 0,25 (na behandeling) (figuur 4C). Deze resultaten wijzen er sterk op dat deze toename van de P700-oxidatieverhouding niet kan worden verklaard door de verandering in pmf, die grotendeels verschilt van de correlatie tussen Y(ND) en pmf onder hoog licht (Yamamoto et al., 2016; Takagi et al., 2017). Daarom is het regulerende effect van PMF op de P700-oxidatieverhouding minimaal bij weinig licht, maar wordt het vooral cruciaal bij veel licht of fluctuerend licht. Bij hoog licht gaat een hoge waarde van Y(ND) meestal gepaard met een hoge waarde van NPQ (Munekage et al., 2002, 2004; Kono et al., 2014; Zivcak et al., 2015), als gevolg van de toename van pmf. De verzwakking van ΔpH vorming leidt tot verlaagde niveaus van Y (ND) en NPQ onder hoog licht (Suorsa et al., 2012; Kono et al., 2014; Yamamoto et al., 2016; Kanazawa et al., 2017). De inductie van NPQ bij weinig licht wordt voornamelijk bepaald door het niveau van lumenverzuring. Na behandeling met hoog licht veranderde de mate van lumenverzuring niet (figuur 5B) en bleef NPQ stabiel (figuur 4D). Ter vergelijking, y (ND) aanzienlijk toegenomen. Deze resultaten wijzen erop dat bij weinig licht, Y(ND) en NPQ werden gecontroleerd door verschillende regelgevende mechanismen.
Etrii bestuurt P700 Oxidatieverhouding bij weinig licht
PSI wordt alleen over-gereduceerd wanneer de elektronenstroom van PSII de capaciteit van PSI-elektronacceptoren overschrijdt om met de elektronen om te gaan. Wanneer de elektronenstroom naar PSI beperkt is, is PSI extreem tolerant tegen lichte stress in de pgr5-planten van A. thaliana (Suorsa et al., 2012, 2016; Tikkanen et al., 2014). Voor de schaduw-vestigende plant Psychotria rubra, PSI-activiteit was niet waarneembaar voor high-light stress in de aanwezigheid van DCMU (Huang et al., 2016c). In gekoelde bladeren van tabak, matige PSII foto-tentoonstelling toegestaan het behoud van P700 optimaal geoxideerd en vervolgens beschermd PSI-activiteit tegen verdere fotoschade (Huang et al., 2016a). Deze resultaten wezen op de belangrijke rol van ETRII bij het beheersen van de redoxtoestand van PSI onder hoge licht-of koel-lichtspanningen. Onze huidige resultaten toonden aan dat de matige PSII fototentoonstelling leidde tot een significante depressie van ETRII bij weinig licht. Ondertussen veranderden de PSI-activiteit en de activiteit van het chloroplastatp-synthase licht en namen de totale pmf en ΔpH niet toe. Als gevolg hiervan werd de afname van ETRII niet veroorzaakt door de ΔpH-afhankelijke fotosynthetische controle via Cyt B6/f complex, maar waarschijnlijk veroorzaakt door de afname van PSII activiteit (Tikkanen et al., 2014). De verminderde toevoer van elektronen van PSII naar PSI leidde tot het verhoogde niveau van P700-oxidatie. Zo werd de P700 oxidatieverhouding bij weinig licht grotendeels geregeld door ETRII.
conclusie
We vonden dat de selectieve fotoinhibitie van PSII een stimulatie van CEF en een verhoging van de P700 oxidatieverhouding bij weinig licht induceerde. De stimulatie van CEF leidde niet tot de ΔpH-afhankelijke downregulatie van fotosynthetisch elektronentransport. Hierdoor vergemakkelijkt deze stimulatie van CEF bij weinig licht voornamelijk de synthese van ATP, wat essentieel is voor de snelle reparatie van fotobeschadigde PSII. De toename van de P700-oxidatieverhouding kon niet worden verklaard door de verandering in de ΔpH-afhankelijke fotosynthetische controle bij het Cyt b6/f-complex, maar werd voornamelijk veroorzaakt door de verminderde toevoer van elektronen van PSII naar PSI.
Auteursbijdragen
WH, S-BZ en TL ontwierpen en ontwierpen het onderzoek. WH en Y-JY voerden de experimenten uit. WH, Y-JY, S-BZ en TL analyseerden de gegevens. WH schreef het eerste ontwerp van het manuscript, dat door alle auteurs intensief werd bewerkt.
financiering
deze studie werd ondersteund door de National Natural Science Foundation of China (31670343), Youth Innovation Promotion Association of the Chinese Academy of Sciences (2016347), en de grote programma ‘ s Wetenschap en techniek in de provincie Yunnan (2016ZF001).
belangenconflict verklaring
De auteurs verklaren dat het onderzoek werd uitgevoerd zonder enige commerciële of financiële relatie die als een potentieel belangenconflict kon worden opgevat.
Kono, M., Noguchi, K., and Terashima, I. (2014). Rol van de cyclische elektronenstroom rond PSI (CEF-PSI) en O2-afhankelijke alternatieve routes in regulatie van de fotosynthetische elektronenstroom in kortstondig fluctuerend licht in Arabidopsis thaliana. Plantencelfysiol. 55, 990–1004. doi: 10.1093/pcp/pcu033
PubMed Abstract | CrossRef Full Text/Google Scholar
Shikanai, T. (2016). Regulerend netwerk van Proton motive force: bijdrage van cyclisch elektronentransport rond photosystem I. Photosynth. Res. 129, 253-260. doi: 10.1007 / s11120-016-0227-0
PubMed Abstract / CrossRef Full Text / Google Scholar
Yamori, W., Makino, A., and Shikanai, T. (2016). Een fysiologische rol van cyclisch elektronentransport rond fotosysteem I bij het in stand houden van fotosynthese onder fluctuerend licht in rijst. Sci. Rapport 6: 20147. doi: 10.1038/srep20147
PubMed Abstract | CrossRef Full Text/Google Scholar
Yamori, W., and Shikanai, T. (2016). Fysiologische functies van cyclisch elektronentransport rond fotosysteem I in het ondersteunen van fotosynthese en plantengroei. Annu. Rev. Plant Biol. 67, 81–106. doi: 10.1146/annurev-arplant-043015-112002
PubMed Abstract | CrossRef Full Text/Google Scholar
Yamori, W., Shikanai, T., and Makino, A. (2015). Fotosystem I cyclische elektronenstroom via chloroplast NADH dehydrogenase-achtig complex speelt een fysiologische rol bij fotosynthese bij weinig licht. Sci. Rep. 5: 13908. doi: 10.1038/srep13908
PubMed Abstract | CrossRef Full Text/Google Scholar
Zhang, S. P., and Scheller, H. V. (2004). Fototentoonstelling van fotosysteem I bij koelende temperatuur en vervolgens herstel in Arabidopsis thaliana. Plantencelfysiol. 45, 1595–1602. doi: 10.1093/pcp/pch180
PubMed Abstract / CrossRef Full Text / Google Scholar