- Inleiding
- methode
- deelnemers
- beoordeling van creativiteit
- Torrance Test of Creative Thinking (Circles Sub-Scale)
- Alternate Uses Task (AUT)
- Pretest
- DNA-extractie en genotypering
- statistische analyse
- resultaten
- de Fluency Component
- de Flexibiliteitscomponent
- de Originaliteitscomponent
- genderverschillen
- discussie
- belangenconflict verklaring
Inleiding
Creatieve cognitie speelt een belangrijke rol in de Kunsten, in uitvinding en innovatie, evenals in het dagelijks leven (Runco and Richards, 1997). Hoewel creativiteit wordt beschouwd als een unieke menselijke capaciteit, spontaan Creatief gedrag (bijv., creating new tools among primates and birds) is aangetoond dat het ook bij niet-mensen voorkomt (Byrne and Bates, 2007), wat verder getuigt van de diepe evolutionaire en biologische wortels van creativiteit. Creativiteit is gedefinieerd als het vermogen om reacties te produceren die zowel nieuw (d.w.z. origineel, zeldzaam en onverwacht) als geschikt (d.w.z., adaptief en nuttig volgens taakbeperkingen) zijn (Sternberg and Lubart, 1999). Een van de psychometrische benaderingen van het meten van creativiteit omvat divergent thinking (DT) taken waarbij deelnemers worden gevraagd om met meerdere oplossingen op een gegeven probleem te reageren (Dietrich en Kanso, 2010). DT-Tests meten in het algemeen verschillende aspecten van creativiteit, waaronder creatieve vloeiendheid, flexibiliteit en originaliteit (Torrance, 1974). Dus, hoewel niet synoniem met creativiteit, DT taken bieden gestructureerde en objectieve metingen van creativiteit (Sternberg en Lubart, 1999; Jung et al ., 2009). Met name de scores op dt-taken zijn positief gecorreleerd met ecologisch geldige maatstaven van creatieve prestatie (Carson et al., 2005) en met eigen creativiteit (Furnham and Bachtiar, 2008).eerdere studies hebben gewezen op de betrokkenheid van het dopaminerge (DA) systeem bij creativiteit (Heilman et al., 2003; Flaherty, 2005; Takeuchi et al., 2010). Takeuchi et al. (2010) vond individuele verschillen in creativiteit, zoals gemeten door DT, positief gecorreleerd met grijze materie in Da-systeemregio ‘ s, waaronder de dorsolaterale prefrontale cortex, bilaterale basale ganglia, substantia nigra, en het tegmentale ventrale gebied. Bovendien hebben verscheidene genetische studies een verband tussen DT en dopamine neurotransmissie aangetoond (Reuter et al., 2006; de Manzano et al., 2010; Runco et al., 2011). Bijvoorbeeld, Reuter et al. (2006) vond creativiteit, zoals gemeten door DT-taken waarbij zowel figurale als verbale creativiteit betrokken zijn, significant geassocieerd met polymorfismen van het dopamine D2 receptor gen (DRD2). Bovendien hebben verscheidene studies corticale dopamine gevonden om in cognitieve flexibiliteit betrokken te zijn (Frank, 2005; Cools, 2008; Garcia-Garcia et al ., 2010), een van de belangrijkste componenten van DT.
een interessant da-kandidaatgen voor creativiteit is het dopamine D4-receptorgen (DRD4). De DRD4-receptor is één van de vijf dopamine-receptoren en speelt een belangrijke rol in het bemiddelen van het synaptische dopamine signaleren. Het gen wordt gekenmerkt door een 48 base-paar variabel aantal achter elkaar herhalingen (48-bp VNTR) gelegen in het codeergebied van de derde exon. DRD4 48 – BP VNTR polymorfisme varieert van 2 tot 11 herhalingen tussen individuen (Asghari et al., 1995; Cravchik and Goldman, 2000), waar de 4-herhaling (4R) de meest voorkomende herhaling is in Kaukasische populaties en de 7-herhaling (7R) de tweede meest voorkomende variant is (Chang et al., 1996; Ding et al., 2002). Interessant, de 7R is eerder geassocieerd met real-life Creatief gedrag, zoals de nieuwheid-zoekende persoonlijkheidskenmerken (Ebstein et al., 1996). Inderdaad, novelty-seeking – de neiging tot exploratieve activiteit-wordt beschouwd als een van de kenmerken van creatieve mensen (Chavez-Eakle et al., 2006; Drago et al., 2009). Hoewel het bewijs voor een verband tussen DRD4 7R en het zoeken naar nieuwigheid inconsistent is, zoals blijkt uit verschillende meta-analyserapporten (Kluger et al., 2002; Munafò et al., 2008), hebben veel studies een significant verband gevonden tussen het zoeken naar nieuwigheid en het 7R-allel van de DRD4 (Ebstein et al., 1996; Benjamin et al., 2000; Keltikangas-Järvinen et al., 2002; Becker et al., 2005). Roussos et al. (2009) hebben onlangs gesuggereerd dat verschillen in het meten van novelty-seeking met behulp van self-report schalen kunnen verklaren voor de inconsistente resultaten tussen 7R polymorfisme en novelty-seeking. In overeenstemming hiermee is herhaaldelijk vastgesteld dat het DA-systeem een belangrijke rol speelt in de persoonlijkheidskenmerken van het zoeken naar nieuwigheid (Flaherty, 2005; Schweizer, 2006), wat de potentiële rol van DRD4 in creativiteit verder bevestigt.in tegenstelling tot de rol van DRD4 48-bp VNTR 7R bij het zoeken naar nieuwigheden, blijkt uit recent onderzoek dat 7R in feite geassocieerd wordt met verminderde cognitieve flexibiliteit, een van de belangrijkste aspecten van creativiteit (Strobel et al., 2004; Congdon et al., 2008). Flexibiliteit omvat de mogelijkheid om een alternatieve reactie te maken na het succesvol remmen van een huidige reactie. De bevindingen tot nu toe over het verband tussen remming en DRD4 zijn gemengd. Congdon et al. (2008) vond dat deelnemers met de 7R allel van de DRD4 vertonen hogere stop-signaal reactietijd (SSRT) op een go/no-go taak, als gevolg van slechtere remmende controle, terwijl Forbes et al. (2009) een dergelijk effect op de Barratt impulsiviteit Schaal (BIS), een self-report tool om impulsiviteit te meten, niet gevonden. Aan de andere kant, Colzato et al. (2010) toonde aan dat deze gemengde effecten te wijten kunnen zijn aan het feit dat eerdere studies betrekking impulsiviteit als een monolithisch proces, terwijl impulsiviteit daadwerkelijk kan worden onderverdeeld in functionele en disfunctionele types volgens Dickman ‘ s impulsiviteit inventaris (DII; Dickman, 1990). Disfunctionele impulsiviteit is de neiging om te handelen zonder vooruitdenken in gevallen waarin een dergelijke actie is ongepast, terwijl functionele impulsiviteit is een soortgelijke tendens uitgevoerd in geschikte situaties (Colzato et al., 2010). Colzato et al. (2010) vond dat de individuen met DRD4 7R, die met hogere niveaus van striatal DA wordt geassocieerd, hogere disfunctionele impulsiviteit tentoonstelden. Verschillende meta-analytische studies hebben gewezen op de 7R allel van de DRD4 als een risico allel voor aandacht-tekort hyperactiviteit stoornis (ADHD; Faraone et al., 2001; DiMaio et al., 2003), een stoornis gekenmerkt door een hoge disfunctionele impulsiviteit (Young et al., 2007). Verder is de relatie tussen ADHD en creativiteit onderzocht in verschillende studies (Shaw and Brown, 1990; Healey and Rucklidge, 2006), hoewel deze studies tot nu toe onduidelijke resultaten hebben opgeleverd. Terwijl sommigen ADHD en zijn symptomologie positief gecorreleerd hebben met creativiteit (Healey and Rucklidge, 2006; White and Shah, 2011), hebben anderen een tegenovergestelde trend gemeld (Funk et al., 1993; Healey and Rucklidge, 2005, 2008). Interessant, wordt ADHD gedragsmatig geassocieerd met impairments in uitvoerende controle, met inbegrip van attentional set-shifting (Boonstra et al., 2005), en flexibiliteit (Barkley et al., 1997; Sergeant et al., 2002), die beide belangrijk zijn voor cognitieve flexibiliteit, een centrale dimensie van creativiteit (Dietrich, 2004; Durstewitz and Seamans, 2008). Deze bevindingen wijzen erop dat personen met het 7R-allel van de DRD4 een lagere mate van creativiteit en verminderde flexibiliteit kunnen vertonen met betrekking tot DT-taken in het bijzonder. Daarom is de DRD4 exon III VNTR een biologisch plausibele kandidaat om bij te dragen aan individuele verschillen in creativiteit.
gezamenlijk blijkt dat enerzijds het 7R-allel, als risico-allel voor verminderde uitvoerende functies, flexibiliteit en set shifting, geassocieerd kan worden met een lage creativiteit. Aan de andere kant, de associatie tussen de 7R allel en hogere nieuwheid-zoeken geeft aan dat individuen met de 7R allel kan eigenlijk vertonen meer flexibiliteit en creativiteit.
de huidige studie is ontworpen om de complexe rol van DRD4 7R in het gedeeltelijk vormgeven van menselijke creativiteit te onderzoeken. Om deze relatie beter te karakteriseren, beoordeelden we twee soorten DT-tests (figuraal en verbaal). De waardering van elke taak werd verdeeld volgens de drie belangrijkste dimensies van DT, namelijk creatieve vloeiendheid, flexibiliteit en originaliteit, en elke dimensie werd afzonderlijk geanalyseerd om de hypothese te testen dat DRD4 elke dimensie van creativiteit verschillend beïnvloedt.
methode
deelnemers
de steekproef bestond uit 185 studenten van de Universiteit van Haifa in Israël (112 vrouwen). Alle deelnemers waren blank (zelf gemeld), met een gemiddelde leeftijd van 24,5 (SD ± 2,1). Deelnemers werden gerekruteerd door middel van aankondigingen geplaatst op de universiteit. Alle deelnemers waren betaalde vrijwilligers.
beoordeling van creativiteit
creativiteit werd beoordeeld aan de hand van twee DT-tests, een subschaal van de figural sub-test van de Torrance test of creative thinking (Torrance, 1974), en de Alternate Uses Task (Aut; Guilford et al., 1978). Beide tests omvatten de drie kernafmetingen van DT, namelijk flexibiliteit, originaliteit en vloeiendheid.
Torrance Test of Creative Thinking (Circles Sub-Scale)
deelnemers kregen een pagina waarop 30 identieke cirkels werden getekend. Ze werden gevraagd om zoveel mogelijk verschillende betekenisvolle objecten te tekenen binnen een tijdslimiet van 10 minuten, waarbij elke tekening ten minste één cirkel moet bevatten. De scores omvatten vloeiendheid( aantal geproduceerde antwoorden), flexibiliteit (aantal categorieën) en originaliteit, berekend volgens de scores van originele antwoorden, zoals beschreven in de Torrance Tests of Creative Thinking scores guide (Torrance, 1974).
Alternate Uses Task (AUT)
deelnemers kregen een lijst van vijf gemeenschappelijke objecten en werd gevraagd zo veel mogelijk alternatieve toepassingen voor elk object op te sommen, binnen een tijdslimiet van 10 min. Het meest voorkomende dagelijks gebruik werd tussen haakjes aangegeven. De objecten waren: schoen (algemeen gebruik: slijtage op de voet); knop (algemeen gebruik: het sluiten van dingen); potlood (algemeen gebruik: tekenen of schrijven); band (algemeen gebruik: auto wiel); en drinken glas (algemeen gebruik: bevat vloeistof). Alleen reacties die de gegeven algemene toepassingen niet herhaalden werden geteld en opgenomen. Net als in de Torrance-test omvatte scores vloeiendheid, flexibiliteit en originaliteit. Aangezien er geen richtlijnen zijn voor het scoren van originele responsen in de AUT, werden originele responsen gedefinieerd als statistisch infrequente responsen volgens een pretest uitgevoerd in ons lab.
Pretest
met het oog op het creëren van een geldig criterium voor responsfrequentie voltooide een groep van 100 gezonde deelnemers die niet aan dit onderzoek deelnamen de AUT. Voor elk object werd een lijst van alle mogelijke toepassingen verzameld bij alle deelnemers. Op basis van deze lijst werd een statistische meting van de frequentie berekend om de originaliteitsscore voor elk antwoord en vervolgens voor elke deelnemer te evalueren. Antwoorden kregen een score van nul als 5% of meer van de deelnemers vermeld een bepaald gebruik, een score van één als tussen 2% en 4,99% van de deelnemers vermeld het, en een score van twee als minder dan 1,99% vermeld het gebruik. Voor elke deelnemer werd een gemiddelde originaliteitsscore berekend op basis van deze statistische niet-frequente scores.
DNA-extractie en genotypering
DNA werd geëxtraheerd uit 20 ml mondwatermonsters met behulp van de Master Pure kit (Epicentre, Madison, WI). De DRD4 48-bp VNTR werd gekenmerkt door een PCR – versterkingsprocedure met de volgende primers: F5′ – CTT CCT ACC CTG CCC GCT CAT GCTGCT CTA CTGG – 3′ en R5′ – ACC ACCACC GGC AGG ACC CTC ATG GCC TTG CGC TC-3. PCR reacties werden uitgevoerd met 5 µl Master Mix( Thermo scientific), 2 µl primers (0,5 µM), 0,6 µl Mg/Cl2 (2,5 mM), 0.4 µl DMSO 5% en 1 µl water tot een totaal van 9 µl totaal volume, en een extra 1 µl genomisch DNA werd toegevoegd aan het mengsel. Alle PCR-reacties werden uitgevoerd op een Biometra T1 Thermocycler (Biometra, Güttingem, Duitsland). De PCR-reactievoorwaarde was als volgt: voorverwarmingsstap bij 94,0°C gedurende 5 min, 34 denaturatiecycli bij 94,0°C gedurende 30 s, naharden bij 55 ° C gedurende 30 s, en verlenging bij 72°C gedurende 90 s. de reactie bleef 5 min bij 72°C. Het reactiemengsel werd vervolgens elektroforeseerd op een 3% agarose gel (AMRESCO) met ethidiumbromide om genotypes te screenen.
de verdeling van de genotype frequentie was volgens het Hardy-Weinberg evenwicht (chi-kwadraat = 0,01, p-waarde = 0,92).
statistische analyse
Genotype werd geclassificeerd op basis van de aanwezigheid of afwezigheid van het 7R-allel van de DRD4 (7 VS.no7), zoals in overeenstemming met eerdere rapporten in de literatuur (Ding et al., 2002). Groepsverschillen op creativiteitsscores werden statistisch geanalyseerd met ANOVAs, met de aanwezigheid (7) of afwezigheid (no7) van het DRD4 7R-allel als onafhankelijke variabele. Om de verschillende aspecten van DT (vloeiendheid, flexibiliteit, originaliteit) te onderzoeken, hebben we voor elke component een multivariate variantieanalyse (MANOVA) afzonderlijk uitgevoerd.
resultaten
om te bevestigen dat de twee groepen (7, no7) qua leeftijd en opleiding niet significant verschilden, hebben we onafhankelijke t-tests uitgevoerd om de twee groepen te vergelijken (zie Tabel 1 Voor gemiddelden en standaardafwijkingen). Uit deze analyse bleek dat de twee groepen niet verschilden in termen van leeftijd of opleidingsjaren. Niet-parametrische tests (Mann–Whitney) toonden significante verschillen in de frequentieverdeling voor het geslacht aan (Z = 2,11, p < 0,05).
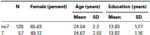
Tabel 1. Leeftijd, geslacht en opleidingsgegevens voor het DRD4 7R genotype, SD: standaarddeviatie.
De gemiddelde scores van de creativiteit die bij de twee taken wordt gebruikt, zijn samengevat in Tabel 2.

Tabel 2. Middelen en standaardfout (SE) van de twee gebruikte creativiteitstaken: AUT en Torrance.
de Fluency Component
een multivariate ANOVA gaf een algemeen effect aan van DRD4 7R genotype voor de fluency component, F(2,182) = 3,25, p < 0,05. Gezien de Betekenis van de algemene test werden de belangrijkste effecten van univariate onderzocht. Significante univariate belangrijkste effecten voor het DRD4-genotype werden verkregen voor AUT-vloeiendheid F(1.183) = 4,95, p < 0.(7R-allel met een lagere vloeibaarheid in vergelijking met de no7-groep, figuur 1A), maar bereikte niet het significantieniveau voor Torrance vloeibaarheid F(1,183) = 2,95, p = 0,087.
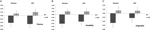
figuur 1. Creativiteit scoort op de aut en Torrance taken gescheiden van vloeiendheid (a), flexibiliteit (B) en originaliteit (C). Middelen werden omgezet in Z-scores.
de Flexibiliteitscomponent
Multivariate ANOVA van de flexibiliteitsmaatregelen vertoonde een algemeen effect voor DRD4 7R genotype F(2,182) = 4.38, p < 0,05. Significante univariate belangrijkste effecten voor DRD4 genotype werden verkregen voor AUT flexibiliteit F(1.183) = 4,57, p < 0,05 en voor Torrance flexibiliteit F(1.183) = 6,95, p < 0,01. Zoals blijkt uit figuur 1B vertoonden in beide gevallen individuen in de 7R-allelgroep minder flexibiliteit dan individuen in de no7-groep (Tabel 2).
de Originaliteitscomponent
Multivariate ANOVA van de originaliteitsmaten vertoonde geen significante effecten, F(2.182) = 2,06, p = 0.13, hoewel de scores wel wijzen op een lagere originaliteit voor het 7R-allel van de DRD4 (zie figuur 1C, Tabel 2 voor details).
genderverschillen
discussie
DT-taken vereisen dat deelnemers meerdere oplossingen bieden voor een bepaald probleem. In de taken die hier werden gebruikt, moesten de deelnemers denken aan vele alternatieve mogelijke toepassingen voor alledaagse items (in de AUT), en om meerdere tekeningen te tekenen met een specifieke vorm (cirkel) (in de Torrance taak). De hier gepresenteerde resultaten tonen aan dat individuen met het DRD4 7R-allel een lagere creativiteit vertonen zoals gemeten door DT. Gezien het feit dat het 7R-allel geassocieerd is met disfunctionele impulsiviteit en slechte remming (Congdon et al., 2008; Dreber et al., 2009), het feit dat individuen met het 7R allel een lagere creativiteit vertonen, kan worden verklaard door hun onvermogen om duidelijke (gewone) reacties te onderdrukken of te remmen, zoals aangegeven door lagere originaliteitsscores. Lagere flexibiliteits-en vloeiendheidsscores kunnen erop wijzen dat het, zodra een gemeenschappelijke respons is gegeven, moeilijker wordt om naar een nieuwe categorie over te schakelen (zoals aangegeven door de lage flexibiliteitsscores), wat leidt tot minder reacties (zoals aangegeven door de lage vloeiendheidsscores). Verder bewijs voor de relatie tussen creativiteit en impulsiviteit kan worden gezien in de associatie van het 7R allel met ADHD (Li et al., 2006), evenals met verhoogde impulsiviteit bij ADHD-patiënten (Langley et al., 2004). Interessant is dat creativiteit is gekoppeld aan het DA-systeem in het geval van schizofrenie (Eysenck, 1993). Hoewel verschillende studies tonen aan dat patiënten met schizofrenie en schizotypische persoonlijkheid vertonen hoge creativiteit (O ‘ Reilly et al., 2001; Folley and Park, 2005) ondersteunen recente modellen van schizofrenie en creativiteit een omgekeerde U-vormige associatie waarin creativiteit hoger kan zijn met een laag-matig schizotypische eigenschap, maar afneemt naarmate de ernst van psychopathologie bij schizofrenie toeneemt (Tsakanikos and Claridge, 2005; Stoneham and Coughtrey, 2009; Nelson and Rawlings, 2010). Inderdaad, in een recente studie Jaracz et al. (2012) rapporteerde dat patiënten met schizofrenie verminderde creativiteit vertonen in vergelijking met controles. Verder rapporteerden de auteurs dat verminderde creativiteit werd geassocieerd met een lage cognitieve flexibiliteit en impulsiviteit, weerspiegeld door hun lage scores op de Wisconsin Card Sorting Test (WCST). Deze resultaten zijn in lijn met eerder onderzoek waaruit blijkt dat patiënten met chronische schizofrenie minder creativiteit vertonen, wat in het geval van creatieve vloeiendheid werd gemedieerd door scores op uitvoerende control taken en in het geval van originaliteit niet was (Abraham et al., 2007). Collectief, terwijl de milde functionele impulsiviteit creativiteit onder patiënten met schizotypal eigenschappen kan verbeteren, vooral in het genereren van ideeën die van de gegeven voorbeelden verschillen (Abraham en Windmann, 2008), kan de dysfunctionele impulsiviteit waargenomen in strenge schizofrenie creatieve vermogens dempen.
de resultaten van de huidige studie ondersteunen de relatie tussen creativiteit en varianten van Da genetische polymorfismen (Reuter et al., 2006; Runco et al., 2011). Bovendien geven de resultaten in onze steekproef aan dat het belangrijkste effect van het DRD4-7R-allel op creativiteit te wijten kan zijn aan de impact ervan op flexibiliteit. Verschillende studies hebben cognitieve flexibiliteit gekoppeld aan da-pathways (Ashby and Isen, 1999; Dreisbach and Goschke, 2004; Müller et al., 2007). Er is gepostuleerd dat de activering van D2-achtige receptoren (d.w.z., D2, D3 en D4) Gabaerge remming vermindert en de activering van veelvoudige voorstellingen vergemakkelijkt, waardoor de flexibiliteit wordt verhoogd die door set-shifting capaciteiten wordt gemanifesteerd (Müller et al., 2007; Seamans and Robbins, 2010). Bovendien zijn D2-achtige receptoren gekoppeld aan flexibele integratie van nieuwe informatie (Durstewitz and Seamans, 2008). In lijn hiermee, Reuter et al. (2006) vond een associatie tussen de dopamine receptor D2 en creativiteit zoals gemeten door een samengestelde index, hoewel niet kan worden bepaald of flexibiliteit een belangrijke rol speelde in deze resultaten. Bovendien, Runco et al. (2011) rapporteerde verschillen in creatieve vloeiendheid tussen dragers van verschillende dopamine genen, waaronder de DRD4. Uit hun verslag blijkt echter niet welke groep meer creatieve vaardigheden bezit. Het is belangrijk op te merken dat in tegenstelling tot de resultaten die hier worden gepresenteerd, Runco et al. (2011) geen verband gevonden tussen flexibiliteitsmaatregelen zoals gemeten door DT-taken en DRD4. Deze inconsistentie kan worden verklaard door verschillen in de gebruikte populaties en de groepsgrootte of door verschillen in allelfrequenties binnen de in de steekproef opgenomen populatie. Verder zijn er aanwijzingen dat de associatie tussen dopamine en DT mogelijk niet lineair is (Akbari Chermahini en Hommel, 2010, 2012), wat erop wijst dat een interactie tussen verschillende da-genen de individuele resultaten van elke genetische variabiliteit in de context van DT kan beïnvloeden. Daarnaast hebben verschillende studies gewezen op het feit dat Da pathways gendergevoelig kunnen zijn (Tammimäki and Männistö, 2011; Wang et al., 2012). Zhong et al. (2010), bijvoorbeeld, vond dat gender gemoduleerd de associatie tussen DRD4 exon3 polymorfisme en gevoel voor eerlijkheid. In de huidige studie hebben we geen interactie-effect gevonden tussen de DRD4-7R en gender, wat aangeeft dat gender geen modulerend effect heeft op de associatie tussen DRD4 en creativiteit in onze steekproef. Bovendien wijzen de hier gepresenteerde resultaten op een gebrek aan genderverschillen in creativiteit, en in DT in het bijzonder (voor een overzicht, zie Baer en Kaufman, 2008).
concluderend, in de huidige studie suggereren we dat de relatie tussen DRD4 en creativiteit voornamelijk wordt beïnvloed door flexibiliteit. De resultaten van de huidige studie zijn in lijn met eerdere resultaten die wijzen op de betrokkenheid van het DA-systeem in creativiteit en toevoegen aan de accumulerende lichaam van kennis suggereren genetische invloeden in creativiteit.
Er zijn verschillende beperkingen van het onderzoek die moeten worden erkend. Ten eerste onderzocht de huidige studie alleen het verband tussen één genetische variabiliteit in het DA—systeem—de DRD4-en DT. Er zijn echter andere belangrijke polymorfismen in het striatale da-systeem die geassocieerd kunnen worden met DT en zowel op een standalone manier als in Gen-geninteracties kunnen bijdragen. Een dergelijk polymorfisme is de dopamine transporter (DAT1), waarvan is aangetoond dat het geassocieerd is met remming van de terugkeer (Colzato et al., 2010) en met maatregelen van impulsiviteit (Gizer and Waldman, 2012). Verder, zoals hierboven vermeld, een andere mogelijkheid die hier niet werd onderzocht is de mogelijke gen-gen interactie effecten op DT. Akbari Chermahini and Hommel (2010) rapporteerden een niet-lineaire associatie tussen een marker van striataal dopamine en DT, wat aangeeft dat de relatie complexer kan zijn, zodanig dat een polymorfisme van een gen invloed kan hebben op DT door de bemiddeling van een ander genpolymorfisme. Daarom moeten toekomstige studies overwegen om de interactie tussen deze twee polymorfismen te onderzoeken in de context van flexibiliteit en DT.
belangenconflict verklaring
De auteurs verklaren dat het onderzoek werd uitgevoerd zonder enige commerciële of financiële relatie die als een potentieel belangenconflict kon worden opgevat.
Akbari Chermahini, S., en Hommel, B. (2010). De (b) link tussen creativiteit en dopamine: spontane knippersnelheden voorspellen en dissociëren afwijkend en convergent denken. Cognitie 115, 458-465. doi: 10.1016 / j. cognitie.2010.03.007
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Cools, R. (2008). Rol van dopamine in de motivatie en cognitieve controle van gedrag. Neurowetenschapper 14, 381-395. doi: 10.1177 / 1073858408317009
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Frank, M. J. (2005). Dynamic dopamine modulation in the basal ganglia: een neurocomputational account of cognitive deficits in medicated and nonmedicated Parkinsonism. J. Cogn. Neurowetenschappen. 17, 51–72. doi: 10.1162 / 0898929052880093
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Healey, D., and Rucklidge, J. J. (2006). Een onderzoek naar de relatie tussen ADHD symptomatologie, creativiteit en neuropsychologisch functioneren bij kinderen. Kinder Neuropsychol. 12, 421–438. doi: 10.1080 / 09297040600806086
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Runco, M. A., and Richards, R. (1997). Eminente creativiteit, alledaagse creativiteit en gezondheid. Greenwich, CT: Ablex.
Torrance, E. (1974). Verschillen zijn geen tekorten. Leren. Coll. Rec. 75, 471–487.