- wprowadzenie
- metoda
- uczestnicy
- ocena kreatywności
- Torrance Test kreatywnego myślenia (Podskala kółek)
- zadanie alternatywne użycia (AUT)
- wstępny test
- ekstrakcja DNA i genotypowanie
- Analiza statystyczna
- wyniki
- Składnik płynnościowy
- Składnik elastyczności
- składowa oryginalności
- różnice płci
- dyskusja
- Oświadczenie o konflikcie interesów
wprowadzenie
kreatywne poznanie odgrywa ważną rolę w sztuce, w wynalazczości i innowacji, a także w życiu codziennym (Runco and Richards, 1997). Choć kreatywność była uważana za wyjątkową zdolność człowieka, spontaniczne zachowania twórcze (np., creating new tools among primates and birds) wykazano, że występują również u Nie-ludzi (Byrne and Bates, 2007), co dodatkowo świadczy o głębokich ewolucyjnych i biologicznych korzeniach kreatywności. Kreatywność została zdefiniowana jako zdolność do tworzenia odpowiedzi, które są zarówno nowatorskie (tj. oryginalne, rzadkie i nieoczekiwane), jak i odpowiednie (tj. adaptacyjne i użyteczne zgodnie z ograniczeniami zadań) (Sternberg i Lubart, 1999). Jednym z psychometrycznych podejść do pomiaru kreatywności są zadania polegające na myśleniu rozbieżnym (divergent thinking, DT), w których uczestnicy są proszeni o odpowiedź na dany problem za pomocą wielu rozwiązań (Dietrich and Kanso, 2010). Testy DT generalnie mierzą różne aspekty kreatywności, w tym kreatywną płynność, elastyczność i oryginalność (Torrance, 1974). Tak więc, chociaż nie jest to synonim kreatywności, zadania DT zapewniają uporządkowane i obiektywne pomiary kreatywności (Sternberg i Lubart, 1999;Jung et al., 2009). W szczególności, wyniki na zadaniach DT okazały się być pozytywnie skorelowane z ekologicznie ważnych środków twórczych osiągnięć (Carson et al., 2005), a także z własną kreatywnością (Furnham i Bachtiar, 2008).
poprzednie badania wskazywały na zaangażowanie układu dopaminergicznego (DA) w kreatywność (Heilman et al., 2003; Flaherty, 2005; Takeuchi et al., 2010). Takeuchi et al. (2010) stwierdził, że indywidualne różnice w kreatywności, mierzone przez DT, są pozytywnie skorelowane z istotą szarą w regionach systemu DA, w tym grzbietowo-bocznej kory przedczołowej, obustronnych zwojów podstawnych, istoty czarnej i nakrywkowej okolicy brzusznej. Dodatkowo, kilka badań genetycznych wykazało związek między DT i neurotransmisji dopaminy (Reuter et al., 2006; De Manzano et al., 2010; Runco et al., 2011). Na przykład, Reuter et al. (2006) stwierdził, że kreatywność, mierzona zadaniami DT obejmującymi zarówno kreatywność figuralną, jak i słowną, jest znacząco związana z polimorfizmami genu receptora dopaminy D2 (DRD2). Ponadto kilka badań wykazało, że dopamina korowa bierze udział w elastyczności poznawczej (Frank, 2005; Cools, 2008; Garcia-Garcia et al., 2010), jeden z głównych elementów DT.
jednym z interesujących genów kandydujących na DA jest gen receptora dopaminy D4 (DRD4). Receptor DRD4 jest jednym z pięciu receptorów dopaminy i odgrywa ważną rolę w mediacji synaptycznej sygnalizacji dopaminy. Gen charakteryzuje się 48-parową zmienną liczbą powtórzeń tandemowych (48-bp VNTR) zlokalizowanych w regionie kodującym trzeci ekson. DRD4 48-bp vntr polimorfizm waha się od 2 do 11 powtórzeń w poszczególnych osobnikach (Asghari et al., 1995; Cravchik and Goldman, 2000), gdzie 4-powtórzenie (4R) jest najczęstszym powtórzeniem w populacjach kaukaskich, a 7-powtórzenie (7R) jest drugim najczęstszym wariantem (Chang et al., 1996; Ding et al., 2002). Co ciekawe, 7R był wcześniej związany z twórczymi zachowaniami w prawdziwym życiu ,takimi jak cecha osobowości poszukującej Nowości(Ebstein et al., 1996). Rzeczywiście, poszukiwanie nowości-tendencja do aktywności eksploracyjnej-jest uważana za jedną z cech kreatywnych ludzi(Chavez-Eakle et al., 2006; Drago et al., 2009). Chociaż dowody na związek między DRD4 7R i poszukiwania nowości są niespójne, o czym świadczą kilka raportów metaanalizy (Kluger et al., 2002; Munafò et al., 2008), wiele badań wykazało znaczący związek między poszukiwaniem nowości a allelem 7R DRD4 (Ebstein et al., 1996; Benjamin et al., 2000; Keltikangas-Järvinen et al., 2002; Becker et al., 2005). Roussos et al. (2009) niedawno zasugerowali, że różnice w pomiarze poszukiwania nowości przy użyciu skal samo-raportowych mogą odpowiadać za niespójne wyniki między polimorfizmem 7R a poszukiwaniem Nowości. W związku z tym wielokrotnie stwierdzono, że system DA odgrywa główną rolę w osobowości poszukiwania nowości (Flaherty, 2005; Schweizer, 2006), co dodatkowo świadczy o potencjalnej roli DRD4 w kreatywności.
niemniej jednak, w przeciwieństwie do roli DRD4 48-bp VNTR 7R w poszukiwaniu nowości, najnowsze dowody wskazują, że 7R jest rzeczywiście związany z upośledzoną elastycznością poznawczą, jednym z głównych aspektów kreatywności (Strobel et al., 2004; Congdon et al., 2008). Elastyczność polega na zdolności do alternatywnej odpowiedzi po pomyślnym zahamowaniu odpowiedzi prądowej. Dotychczasowe wyniki dotyczące związku pomiędzy hamowaniem a DRD4 są mieszane. Congdon et al. (2008) okazało się, że uczestnicy z allelem 7R DRD4 wykazują wyższy czas reakcji sygnału stop (SSRT)na zadanie go / no-go, odzwierciedlając gorszą kontrolę hamującą, podczas gdy Forbes et al. (2009) nie udało się znaleźć takiego wpływu na skalę impulsywności Barratta (BIS), narzędzie do samodzielnego raportowania do pomiaru impulsywności. Z drugiej strony, Colzato et al. (2010) wykazał, że te mieszane efekty mogą być spowodowane faktem, że poprzednie badania dotyczyły impulsywności jako procesu monolitycznego, podczas gdy impulsywność może być faktycznie podzielona na typy funkcjonalne i dysfunkcyjne zgodnie z Dickman ’ s Impulsivity Inventory (Dii; Dickman, 1990). Impulsywność dysfunkcyjna jest tendencją do działania bez przezorności w przypadkach, gdy takie działanie jest niewłaściwe, podczas gdy impulsywność funkcjonalna jest podobną tendencją realizowaną w odpowiednich sytuacjach (Colzato et al., 2010). Colzato et al. (2010) stwierdził, że osoby z DRD4 7R, który jest związany z wyższymi poziomami prążkowia DA, wykazywały wyższą dysfunkcyjną impulsywność. Kilka badań metaanalitycznych wskazało na allel 7R DRD4 jako allel ryzyka dla zespołu nadpobudliwości psychoruchowej z deficytem uwagi(ADHD; Faraone et al., 2001; DiMaio et al., 2003), zaburzenie charakteryzujące się wysoką impulsywnością dysfunkcyjną (Young et al., 2007). Ponadto związek między ADHD a kreatywnością został zbadany w kilku badaniach (Shaw and Brown, 1990; Healey and Rucklidge, 2006), choć do tej pory badania te przyniosły niejasne wyniki. Podczas gdy niektórzy stwierdzili, że ADHD i jego symptomologia są pozytywnie skorelowane z kreatywnością (Healey and Rucklidge, 2006; White and Shah, 2011), inni odnotowali przeciwny trend (Funk et al., 1993; Healey and Rucklidge, 2005, 2008). Co ciekawe, ADHD jest behawioralnie związane z upośledzeniem w kontroli wykonawczej, w tym attentional set-shifting (Boonstra et al., 2005) i elastyczność (Barkley et al., 1997; Sergeant et al., 2002), które są ważne dla elastyczności poznawczej, centralnego wymiaru kreatywności (Dietrich, 2004; Durstewitz and Seamans, 2008). Wyniki te wskazują, że osoby z allelem 7R DRD4 mogą wykazywać niższy poziom kreatywności i zmniejszoną elastyczność w zadaniach DT w szczególności. Stąd DRD4 exon III VNTR jest biologicznie wiarygodnym kandydatem do przyczynienia się do indywidualnych różnic w kreatywności.
łącznie wydaje się, że z jednej strony allel 7R, jako allel ryzyka dla upośledzonych funkcji wykonawczych, elastyczności i przesunięcia zestawu, może być związany z niską kreatywnością. Z drugiej strony związek między allelem 7R a wyższym poszukiwaniem Nowości wskazuje, że osoby z allelem 7R mogą faktycznie wykazywać większą elastyczność i kreatywność.
obecne badanie miało na celu zbadanie złożonej roli DRD4 7R w częściowym kształtowaniu ludzkiej kreatywności. Aby lepiej scharakteryzować tę zależność, oceniliśmy dwa rodzaje testów DT (figuralny i słowny). Punktacja każdego zadania została podzielona zgodnie z trzema głównymi wymiarami DT, a mianowicie płynnością twórczą, elastycznością i oryginalnością, a każdy wymiar był analizowany oddzielnie, aby przetestować hipotezę, że DRD4 w różny sposób wpływa na każdy wymiar kreatywności.
metoda
uczestnicy
w próbie wzięło udział 185 Studentów Uniwersytetu Hajfy w Izraelu (112 kobiet). Wszyscy uczestnicy byli rasy białej (zgłoszeni samodzielnie), średnia wieku wynosiła 24,5 (SD ± 2,1). Uczestników rekrutowano za pośrednictwem ogłoszeń zamieszczonych na Uniwersytecie. Wszyscy uczestnicy byli płatnymi wolontariuszami.
ocena kreatywności
kreatywność oceniano za pomocą dwóch testów DT, podskali z podskali figuralnej testu Torrance ’ a testu kreatywnego myślenia (Torrance, 1974) i alternatywnego zadania zastosowania (AUT; Guilford et al., 1978). Oba testy obejmowały trzy podstawowe wymiary DT, a mianowicie elastyczność, oryginalność i płynność.
Torrance Test kreatywnego myślenia (Podskala kółek)
uczestnicy otrzymali stronę, na której narysowano 30 identycznych kółek. Zostali poproszeni o narysowanie jak największej liczby różnych znaczących obiektów w czasie 10 min, gdzie każdy rysunek musi zawierać co najmniej jeden okrąg. Punktacja obejmowała płynność (liczba odpowiedzi), elastyczność (liczba kategorii) i oryginalność, obliczoną zgodnie z punktacją oryginalnych odpowiedzi, jak opisano w Torrance Tests of Creative Thinking scoring guide (Torrance, 1974).
zadanie alternatywne użycia (AUT)
uczestnicy otrzymali listę pięciu wspólnych obiektów i zostali poproszeni o listę jak największej liczby alternatywnych zastosowań dla każdego obiektu w czasie 10 minut. W nawiasie podano najczęstsze codzienne użycie. Przedmiotami były: but (powszechne zastosowanie: nosić na piechotę); przycisk (wspólne zastosowanie: zamykanie rzeczy); ołówek (wspólne zastosowanie: rysowanie lub pisanie); opona (wspólne zastosowanie: koło samochodu); i szklanka do picia (wspólne zastosowanie: zawiera płyn). Uwzględniono tylko odpowiedzi, które nie powtórzyły danych powszechnych zastosowań. Podobnie jak w teście Torrance, punktacja obejmowała płynność, elastyczność i oryginalność. Ponieważ nie ma wytycznych dotyczących punktacji oryginalnych odpowiedzi w AUT, oryginalne odpowiedzi zostały zdefiniowane jako statystycznie rzadkie odpowiedzi zgodnie z testem wstępnym przeprowadzonym w naszym laboratorium.
wstępny test
w celu stworzenia ważnego kryterium częstości odpowiedzi, Grupa 100 zdrowych uczestników, którzy nie wzięli udziału w tym badaniu, ukończyła AUT. Dla każdego obiektu zebrano listę wszystkich możliwych zastosowań od wszystkich uczestników. Statystyczna miara Rzadkości została obliczona na podstawie tej listy w celu oceny wyniku oryginalności dla każdej odpowiedzi, a następnie dla każdego uczestnika. Odpowiedziom przypisano wynik zerowy, Jeśli 5% lub więcej uczestników wymieniło dane użycie, wynik jeden, jeśli między 2% a 4,99% uczestników wymieniło je, a wynik dwa, jeśli mniej niż 1,99% wymieniło użycie. Średni wynik oryginalności został obliczony dla każdego uczestnika zgodnie z tymi statystycznymi wynikami Rzadkości.
ekstrakcja DNA i genotypowanie
DNA ekstrahowano z 20 ml próbek płynu do płukania jamy ustnej przy użyciu Master Pure kit (Epicentre, Madison, WI). DRD4 48-bp VNTR charakteryzował się procedurą amplifikacji PCR z następującymi starterami: F5′ – CTT CCT ACC CTG CCC GCT CAT GCT GCTGCT CTA CTGG – 3′ i R5 ’ – ACC ACCACC GGC AGG ACC CTC ATG GCC TTG CGC TC-3. Reakcje PCR prowadzono przy użyciu 5 µl Master Mix (Thermo scientific), 2 µl starterów (0,5 µM), 0,6 µl Mg/Cl2 (2,5 mM), 0.Do mieszaniny dodano 4 µl DMSO 5% I 1 µl wody do całkowitej objętości 9 µl i dodatkowy 1 µl genomowego DNA. Wszystkie reakcje PCR przeprowadzono na termocyklerze Biometra T1 (Biometra, Güttingem, Niemcy). Warunki reakcji PCR były następujące: etap wstępnego podgrzewania w temperaturze 94,0 ° C przez 5 min, 34 cykle denaturacji w temperaturze 94,0°C przez 30 s, ponowne podgrzewanie w temperaturze 55°C przez 30 s i przedłużanie w temperaturze 72°C przez 90 s. reakcja utrzymywała się w temperaturze 72°C przez 5 min. Mieszaninę reakcyjną następnie poddano elektroforezie na 3% żelu agarozowym (AMRESCO) z bromkiem etydyny w celu zbadania genotypów.
rozkład częstości genotypu był zgodny z równowagą Hardy ’ ego-Weinberga (chi-kwadrat = 0,01, wartość p = 0,92).
Analiza statystyczna
genotyp został sklasyfikowany na podstawie obecności lub braku allelu 7R DRD4 (7 vs.no7), zgodnie z wcześniejszymi doniesieniami w literaturze (Ding et al., 2002). Różnice grupowe w skali kreatywności analizowano statystycznie przy użyciu ANOVAs, z obecnością (7) lub nieobecnością (no7) allelu DRD4 7R jako niezależną zmienną. Aby zbadać różne aspekty DT (płynność, elastyczność, oryginalność), przeprowadziliśmy wielowymiarową analizę wariancji (MANOVA) oddzielnie dla każdego komponentu.
wyniki
aby potwierdzić, że obie grupy (7, nr 7) nie różniły się znacząco pod względem wieku i wykształcenia, przeprowadziliśmy niezależne testy t w celu porównania obu grup (średnie i odchylenia standardowe patrz Tabela 1). Analiza ta wykazała, że obie grupy nie różniły się pod względem wieku lub lat edukacji. Testy nieparametryczne (Mann–Whitney) wykazały istotne różnice w rozkładzie częstości występowania dla płci (Z = 2,11, p < 0,05).
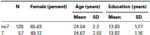
Tabela 1. Wiek, płeć i wykształcenie dane dla genotypu DRD4 7R, SD: odchylenie standardowe.
średnie wyniki środków kreatywności stosowanych w obu zadaniach podsumowano w tabeli 2.

Tabela 2. Średnia i błąd standardowy (SE) dwóch użytych zadań kreatywnych: AUT i Torrance.
Składnik płynnościowy
wielowymiarowa ANOVA wskazywała na ogólny efekt genotypu DRD4 7R dla składnika płynnościowego, F(2,182) = 3,25, p< 0,05. Biorąc pod uwagę znaczenie całego testu, zbadano jednorodne główne efekty. Znaczące, jednowymiarowe główne efekty dla genotypu DRD4 uzyskano dla płynności AUT F(1,183) = 4,95, p < 0.05 (allel 7R wykazujący mniejszą płynność w porównaniu z grupą no7, Fig.1a), ale nie osiągnął poziomu istotności dla płynności Torrance ’ A F(1183) = 2,95, P = 0,087.
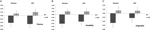
Rysunek 1. Kreatywność odnosi się do zadań AUT i Torrance rozdzielonych na płynność (a), elastyczność (B) i oryginalność (C). Środki zostały przekształcone w wyniki Z.
Składnik elastyczności
wielowymiarowa ANOVA miar elastyczności wykazywała ogólny efekt dla genotypu DRD4 7R F(2182) = 4.38, p < 0.05. Istotne jednowymiarowe główne efekty dla genotypu DRD4 uzyskano dla elastyczności AUT F(1,183) = 4,57, p < 0,05 i dla elastyczności Torrance ’ A F(1,183) = 6,95, p < 0,01. Jak pokazano na fig. 1B, w obu przypadkach osoby z grupy alleli 7R wykazywały mniejszą elastyczność w porównaniu z osobami z grupy no7(Tabela 2).
składowa oryginalności
Wielowymiarowa ANOVA miar oryginalności nie wykazywała żadnych skutków istotnych, F(2182) = 2,06, p = 0.13, chociaż wyniki wskazywały na mniejszą oryginalność allelu 7R DRD4 (szczegóły patrz rysunek 1C, Tabela 2).
różnice płci
dyskusja
zadania DT wymagają od uczestników dostarczenia wielu rozwiązań danego problemu. W zastosowanych tu zadaniach uczestnicy musieli wymyślić wiele alternatywnych możliwych zastosowań przedmiotów codziennego użytku (w AUT) i narysować wiele rysunków zawierających określony Kształt (Okrąg) (w zadaniu Torrance). Przedstawione tutaj Wyniki pokazują, że osoby z allelem DRD4 7R wykazują mniejszą kreatywność mierzoną przez DT. Biorąc pod uwagę, że allel 7R jest związany z dysfunkcyjną impulsywnością i słabym hamowaniem (Congdon et al., 2008; Dreber et al., 2009), fakt, że osoby z allelem 7R wykazują niższą kreatywność, można wyjaśnić ich niezdolnością do tłumienia lub hamowania oczywistych (powszechnych) odpowiedzi, na co wskazują niższe wyniki oryginalności. Niższa ocena elastyczności i płynności może wskazywać, że po udzieleniu wspólnej odpowiedzi trudniej jest przejść do nowej kategorii (na co wskazują niskie wyniki elastyczności), co prowadzi do mniejszej liczby odpowiedzi (na co wskazuje niski wynik płynności). Dalsze dowody na związek między kreatywnością a impulsywnością można zobaczyć w związku allelu 7R z ADHD (Li et al., 2006), jak również ze zwiększoną impulsywnością u pacjentów z ADHD (Langley et al., 2004). Co ciekawe, kreatywność została powiązana z systemem DA w przypadku schizofrenii (Eysenck, 1993). Chociaż kilka badań pokazuje, że pacjenci ze schizofrenią i schizotypową osobowością wykazują wysoką kreatywność (O ’ Reilly et al., 2001; Folley and Park, 2005), recent models of schizophrenia and creativity support an inverted U-shaped association in which creativity may be higher with low-moderate schizotypal trait, but maleje as the severity of psychopathology in schizophrenia increases (Tsakanikos and Claridge, 2005; Stoneham and Coughtrey, 2009; Nelson and Rawlings, 2010). Rzeczywiście, w niedawnym badaniu Jaracz et al. (2012) poinformował, że pacjenci ze schizofrenią wykazują zmniejszoną kreatywność w porównaniu do kontroli. Ponadto autorzy stwierdzili, że upośledzenie kreatywności wiązało się z niską elastycznością poznawczą i impulsywnością, co znalazło odzwierciedlenie w ich niskich wynikach w Wisconsin Card Sorting Test (WCST). Wyniki te są zgodne z wcześniejszymi badaniami pokazującymi, że pacjenci z przewlekłą schizofrenią wykazują niższą kreatywność, która w przypadku płynności twórczej była pośredniczona przez Wyniki na zadaniach kontroli wykonawczej, a w przypadku oryginalności nie była (Abraham et al., 2007). Łącznie, podczas gdy łagodna impulsywność czynnościowa może poprawić kreatywność u pacjentów z cechami schizotypowymi, zwłaszcza w generowaniu pomysłów, które różnią się od podanych przykładów (Abraham and Windmann, 2008), dysfunkcyjna impulsywność obserwowana w ciężkiej schizofrenii może tłumić zdolności twórcze.
wyniki obecnego badania potwierdzają związek między kreatywnością a wariantami polimorfizmów genetycznych DA (Reuter et al., 2006; Runco et al., 2011). Co więcej, wyniki Znalezione w naszej próbie wskazują, że główny wpływ allelu DRD4-7R na kreatywność może wynikać z jego wpływu na elastyczność. Kilka badań powiązało elastyczność poznawczą ze ścieżkami DA (Ashby and Isen, 1999; Dreisbach and Goschke, 2004; Müller et al., 2007). Postulowano, że aktywacja receptorów podobnych do D2 (tj. D2, D3 i D4) zmniejsza hamowanie GABAergiczne i ułatwia aktywację wielu reprezentacji, zwiększając w ten sposób elastyczność przejawiającą się zdolnościami zmiany ustawień (Müller et al., 2007; Seamans and Robbins, 2010). Ponadto receptory D2-podobne zostały połączone z elastyczną integracją nowych informacji (Durstewitz i Seamans, 2008). Zgodnie z tym, Reuter et al. (2006) stwierdził związek między receptorem dopaminy D2 a kreatywnością mierzoną przez złożony indeks, chociaż nie można określić, czy elastyczność odegrała główną rolę w tych wynikach. Dodatkowo, Runco et al. (2011) odnotowano różnice w płynności twórczej wśród nosicieli kilku genów dopaminy, wśród nich DRD4. Z ich raportu nie wynika jednak jasno, która grupa wykazuje większe zdolności twórcze. Ważne jest, aby pamiętać, że w przeciwieństwie do wyników przedstawionych tutaj, Runco et al. (2011) nie znaleziono związku między miarami elastyczności mierzonymi zadaniami DT a DRD4. Niespójność tę można wyjaśnić różnicami w zastosowanych populacjach i wielkościach grup lub różnicami w częstości występowania alleli w populacji objętej próbą. Ponadto istnieją dowody sugerujące, że związek między dopaminą a DT może nie być liniowy (Akbari Chermahini i Hommel, 2010, 2012), co wskazuje, że interakcja między kilkoma genami DA może wpływać na indywidualne wyniki każdej zmienności genetycznej w kontekście DT. Ponadto kilka badań wskazuje na fakt, że ścieżki DA mogą być wrażliwe na płeć (Tammimäki and Männistö, 2011; Wang et al., 2012). Zhong i in. (2010), na przykład, stwierdził, że płeć modulowała związek między polimorfizmem DRD4 exon3 a poczuciem sprawiedliwości. W obecnym badaniu nie znaleźliśmy efektu interakcji między DRD4-7R a płcią, co wskazuje, że płeć nie ma wpływu modulującego na związek między DRD4 a kreatywnością w naszej próbie. Ponadto przedstawione tutaj wyniki wskazują na brak różnic między płciami w kreatywności, a w szczególności w DT (przegląd, patrz Baer and Kaufman, 2008).
podsumowując, w obecnym badaniu sugerujemy, że związek między DRD4 a kreatywnością zależy głównie od elastyczności. Wyniki obecnego badania są zgodne z wcześniejszymi wynikami wskazującymi na zaangażowanie systemu DA w kreatywność i uzupełniają gromadzącą się wiedzę sugerującą wpływy genetyczne w kreatywność.
istnieje kilka ograniczeń badania, które należy uznać. Po pierwsze, w obecnym badaniu zbadano tylko związek między jedną zmiennością genetyczną w systemie DA-DRD4-i DT. Istnieją jednak inne ważne polimorfizmy w układzie prążkowia DA, które mogą być związane z DT i mogą przyczyniać się zarówno w sposób samodzielny, jak i w interakcjach Gen-Gen. Jednym z takich polimorfizmów jest transporter dopaminy (DAT1), który, jak wykazano, jest związany z hamowaniem powrotu (Colzato et al., 2010), a także z miarą impulsywności (Gizer i Waldman, 2012). Ponadto, jak wspomniano powyżej, inną możliwością, która nie została zbadana tutaj, jest możliwy wpływ interakcji Gen-Gen na DT. Akbari Chermahini i Hommel (2010) zgłaszali nieliniowy związek między markerem dopaminy w prążkowiu a DT, wskazując, że związek może być bardziej złożony, tak że jeden polimorfizm genu może wpływać na DT poprzez mediację innego polimorfizmu genu. Dlatego przyszłe badania powinny rozważyć zbadanie interakcji między tymi dwoma polimorfizmami w kontekście elastyczności i DT.
Oświadczenie o konflikcie interesów
autorzy oświadczają, że badanie zostało przeprowadzone przy braku jakichkolwiek relacji handlowych lub finansowych, które mogłyby być interpretowane jako potencjalny konflikt interesów.
Akbari Chermahini, S., and Hommel, B. (2010). (B) związek między kreatywnością a dopaminą: spontaniczne mruganie oczu przewiduje i dysocjuje rozbieżne i zbieżne myślenie. Poznań 115, 458-465. doi: 10.1016 / j.Poznań.2010.03.007
PubMed Streszczenie / PubMed Pełny tekst/CrossRef Pełny tekst
Rola dopaminy w motywacyjnej i poznawczej kontroli zachowania. Neurologist 14, 381-395. doi: 10.1177/1073858408317009
PubMed Streszczenie | PubMed Pełny tekst/CrossRef Pełny tekst
Frank, M. J. (2005). Dynamic dopamina modulation in the basal ganglia: a neurocomputational account of cognitive deficits in medicated and non-medicated parkinsonizm. J. Cogn. Neurosci. 17, 51–72. doi: 10.1162/0898929052880093
PubMed Streszczenie | PubMed Pełny tekst/CrossRef Pełny tekst
Healey, D., and Rucklidge, J. J. (2006). Badanie związku między symptomatologią ADHD, kreatywnością i funkcjonowaniem neuropsychologicznym u dzieci. Neuropsychol Dziecięcy. 12, 421–438. doi: 10.1080/09297040600806086
PubMed Streszczenie | PubMed Pełny tekst/CrossRef Pełny tekst
Runco, M. A., and Richards, R. (1997). Wybitna kreatywność, codzienna kreatywność i zdrowie. Greenwich, CT: Ablex.
Różnice nie są deficytami. Naucz. Coll. Rec. 75, 471–487.