- introduktion
- metode
- deltagere
- vurdering af kreativitet
- Torrance Test of Creative Thinking (Circles Sub-Scale)
- alternativ anvendelse opgave (AUT)
- Pretest
- DNA-ekstraktion og genotypebestemmelse
- statistisk analyse
- resultater
- flydende komponent
- Fleksibilitetskomponenten
- Originalitetskomponenten
- kønsforskelle
- Diskussion
- interessekonflikt Erklæring
introduktion
kreativ kognition spiller en vigtig rolle inden for kunst, opfindelse og innovation såvel som i hverdagen (Runco og Richards, 1997). Selvom kreativitet er blevet betragtet som en unik menneskelig kapacitet, spontant kreativ adfærd (f. eks., at skabe nye værktøjer blandt primater og fugle) har vist sig at forekomme hos ikke-mennesker også (Byrne og Bates, 2007), hvilket yderligere vidner om kreativitetens dybe evolutionære og biologiske rødder. Kreativitet er defineret som evnen til at producere svar, der både er nye (dvs.originale, sjældne og uventede) og egnede (dvs. adaptive og nyttige i henhold til opgavebegrænsninger) (Sternberg og Lubart, 1999). En af de psykometriske tilgange til måling af kreativitet involverer divergent thinking (DT) opgaver, hvor deltagerne bliver bedt om at reagere på et givet problem med flere løsninger (Dietrich og Kanso, 2010). Test af DT måler generelt forskellige aspekter af kreativitet, herunder kreativ flydende, fleksibilitet og originalitet (Torrance, 1974). Således, selvom det ikke er synonymt med kreativitet, giver dt-opgaver strukturerede og objektive målinger af kreativitet (Sternberg og Lubart, 1999; Jung et al., 2009). Især har scoringer på DT-opgaver vist sig at være positivt korreleret med økologisk gyldige mål for kreativ præstation (Carson et al., 2005) såvel som med selvvurderet kreativitet (Furnham og Bachtiar, 2008).
tidligere undersøgelser har peget på inddragelsen af det dopaminerge (da) system i kreativitet (Heilman et al., 2003; Flaherty, 2005; Takeuchi et al., 2010). Takeuchi et al. (2010) fandt, at individuelle forskelle i kreativitet, målt ved DT, var positivt korreleret med gråt stof i DA-systemregioner, herunder den dorsolaterale præfrontale bark, bilaterale basalganglier, substantia nigra og det tegmentale ventrale område. Derudover har flere genetiske undersøgelser vist et forhold mellem DT og dopamin neurotransmission (Reuter et al., 2006; De Mansano et al., 2010; Runco et al., 2011). For eksempel Reuter et al. (2006) fandt, at kreativitet målt ved dt-opgaver, der involverede både figur-og verbal kreativitet, var signifikant forbundet med polymorfier af dopamin D2-receptorgenet (DRD2). Desuden har flere undersøgelser fundet, at kortikal dopamin er involveret i kognitiv fleksibilitet (Frank, 2005; Cools, 2008; Garcia-Garcia et al., 2010), en af hovedkomponenterne i DT.
et interessant da-kandidatgen for kreativitet er dopamin D4-receptorgenet (DRD4). DRD4-receptoren er en af fem dopaminreceptorer og spiller en vigtig rolle i mediering af synaptisk dopaminsignalering. Genet er karakteriseret ved et 48 basepar variabelt antal tandem-gentagelser (48-bp VNTR) placeret i kodningsområdet for den tredje ekson. DRD4 48-bp VNTR polymorfisme varierer fra 2 til 11 gentagelser på tværs af individer (Asghari et al., 1995; Cravchik og Goldman, 2000), hvor 4-gentagelsen (4R) er den mest almindelige gentagelse i kaukasiske populationer, og 7-gentagelsen (7R) er den næst mest almindelige variant (Chang et al., 1996; Ding et al., 2002). Interessant nok har 7R tidligere været forbundet med virkelige kreative adfærd, såsom den nyhedssøgende personlighedstræk (Ebstein et al., 1996). Ja, nyhedssøgning—tendensen til sonderende aktivitet-menes at være et af kendetegnene for kreative mennesker (Chaves-Eakle et al., 2006; Drago et al., 2009). Selvom beviset for en sammenhæng mellem DRD4 7R og nyhedssøgning er inkonsekvent, som det fremgår af flere metaanalyserapporter (Kluger et al., 2002; Munaf Kristian et al., 2008), har mange undersøgelser fundet en signifikant sammenhæng mellem nyhedssøgning og 7R-allelen af DRD4 (Ebstein et al., 1996; Benjamin et al., 2000; Keltikangas-J Kurrvinen et al., 2002; Becker et al., 2005). Roussos et al. (2009) har for nylig antydet, at forskelle i måling af nyhedssøgning ved hjælp af selvrapporteringsskalaer kan redegøre for de inkonsekvente resultater mellem 7R-polymorfisme og nyhedssøgning. I tråd med dette er det gentagne gange blevet konstateret, at DA-systemet spiller en vigtig rolle i personlighedstræk ved nyhedssøgning (Flaherty, 2005, 2006), hvilket yderligere vidner om DRD4 ‘ s potentielle rolle i kreativitet.
ikke desto mindre, i modsætning til rollen som DRD4 48-bp VNTR 7R i nyhedssøgning, viser nylige beviser, at 7R faktisk er forbundet med nedsat kognitiv fleksibilitet, et af de vigtigste aspekter af kreativitet (Strobel et al., 2004; Congdon et al., 2008). Fleksibilitet indebærer evnen til at lave et alternativt svar efter vellykket hæmning af et aktuelt svar. Fund hidtil på sammenhængen mellem hæmning og DRD4 er blandet. Congdon et al. (2008) fandt ud af, at deltagere med 7R-allelen i DRD4 udviser højere stop-signalreaktionstid (ssrt) på en go/no-go-opgave, hvilket afspejler dårligere hæmmende kontrol, mens Forbes et al. (2009) kunne ikke finde en sådan effekt på Barratt Impulsiveness Scale (BIS), et selvrapporteringsværktøj til måling af impulsivitet. På den anden side, Coltsato et al. (2010) viste, at disse blandede effekter kan skyldes, at tidligere undersøgelser behandlede impulsivitet som en monolitisk proces, mens impulsivitet faktisk kan opdeles i funktionelle og dysfunktionelle typer i henhold til Dickmans Impulsivitetsopgørelse (DII; Dickman, 1990). Dysfunktionel impulsivitet er tendensen til at handle uden omtanke i tilfælde, hvor en sådan handling er upassende, mens funktionel impulsivitet er en lignende tendens implementeret i passende situationer., 2010). Et al. (2010) fandt ud af, at personer med DRD4 7R, som er forbundet med højere niveauer af striatal da, udviste højere dysfunktionel impulsivitet. Flere metaanalytiske undersøgelser har peget på 7R-allelen af DRD4 som en risikoallel for opmærksomhedsunderskud hyperaktivitetsforstyrrelse (ADHD; Faraone et al., 2001; DiMaio et al., 2003), en lidelse præget af høj dysfunktionel impulsivitet (Young et al., 2007). Desuden er forholdet mellem ADHD og kreativitet blevet undersøgt i flere undersøgelser (1990; Healey og Rucklidge, 2006), selvom disse undersøgelser hidtil har givet uklare resultater. Mens nogle har fundet ADHD og dets symptomologi at være positivt korreleret med kreativitet (Healey og Rucklidge, 2006; hvid og Shah, 2011), har andre rapporteret en modsat tendens (Funk et al., 1993; Healey og Rucklidge, 2005, 2008). Interessant nok er ADHD adfærdsmæssigt forbundet med svækkelser i udøvende kontrol, herunder opmærksom sætskift (Boonstra et al., 2005), og fleksibilitet (Barkley et al., 1997; Sergent et al., 2002), som begge er vigtige for kognitiv fleksibilitet, en central dimension af kreativitet (Dietrich, 2004; Seamans, 2008). Disse fund indikerer, at personer med 7R-allelen i DRD4 kan udvise lavere niveauer af kreativitet og nedsat fleksibilitet på især dt-opgaver. Derfor er DRD4 ekson III VNTR en biologisk plausibel kandidat til at bidrage til individuelle forskelle i kreativitet.
samlet ser det ud til, at 7R-allelen på den ene side som en risikoallel for nedsatte udøvende funktioner, Fleksibilitet og sætskift kan være forbundet med lav kreativitet. På den anden side indikerer sammenhængen mellem 7R-allelen og højere nyhedssøgning, at personer med 7R-allelen faktisk kan udvise større fleksibilitet og kreativitet.
den aktuelle undersøgelse var designet til at udforske DRD4 7R ‘ s komplekse rolle i delvis udformning af menneskelig kreativitet. For bedre at karakterisere dette forhold vurderede vi to typer DT-test (figur og verbal). Scoringen af hver opgave blev opdelt efter de tre hoveddimensioner af DT, nemlig kreativ flydende, fleksibilitet og originalitet, og hver dimension blev analyseret separat for at teste hypotesen om, at DRD4 differentielt påvirker hver dimension af kreativitet.
metode
deltagere
prøven omfattede 185 studerende fra University of Haifa i Israel (112 kvinder). Alle deltagere var kaukasiske (selvrapporterede) med en gennemsnitlig alder på 24,5 (SD-kr.2,1). Deltagerne blev rekrutteret gennem meddelelser, der blev sendt på universitetet. Alle deltagere blev betalt frivillige.
vurdering af kreativitet
kreativitet blev vurderet ved hjælp af to dt-tests, en underskala fra den figurative undertest af Torrance test of creative thinking (Torrance, 1974), og den alternative bruger opgave (aut; Guilford et al., 1978). Begge tests omfattede de tre kernedimensioner af DT, nemlig fleksibilitet, originalitet og flydende.
Torrance Test of Creative Thinking (Circles Sub-Scale)
deltagerne fik en side, hvor 30 identiske cirkler blev tegnet. De blev bedt om at tegne så mange forskellige meningsfulde objekter som muligt inden for en frist på 10 min, hvor hver tegning skal indeholde mindst en cirkel. Scoring inkluderet flydende (antal producerede svar), fleksibilitet (antal kategorier) og originalitet, beregnet i henhold til scoringen af originale svar, som beskrevet i Torrance test of Creative Thinking scoring guide (Torrance, 1974).
alternativ anvendelse opgave (AUT)
deltagerne fik en liste over fem almindelige objekter og blev bedt om at liste så mange alternative anvendelser som muligt for hvert objekt inden for en frist på 10 min. Den mest almindelige daglige brug blev angivet i parentes. Objekterne var: sko (almindelig brug: slid til fods); knap (almindelig brug: lukning af ting); blyant (almindelig brug: tegning eller skrivning); dæk (almindelig brug: bilhjul); og drikkeglas (almindelig brug: indeholder væske). Kun svar, der ikke gentog de givne almindelige anvendelser, blev talt og inkluderet. Som i Torrance-testen inkluderede scoring flydende, fleksibilitet og originalitet. Da der ikke er nogen retningslinjer for scoring af originale svar i AUT, originale svar blev defineret som statistisk sjældne svar i henhold til en prætest udført i vores laboratorium.
Pretest
med det formål at skabe et gyldigt kriterium for responsfrekvens afsluttede en gruppe på 100 raske deltagere, der ikke deltog i denne undersøgelse, AUT. For hvert objekt blev der samlet en liste over alle mulige anvendelser fra alle deltagere. En statistisk sjældne foranstaltning blev beregnet på baggrund af denne liste for at evaluere originalitetsscore for hvert svar og derefter for hver deltager. Svarene blev tildelt en score på nul, hvis 5% eller mere af deltagerne anførte en given anvendelse, en score på en, hvis mellem 2% og 4,99% af deltagerne anførte den, og en score på to, hvis mindre end 1,99% angav brugen. En gennemsnitlig originalitetsscore blev beregnet for hver deltager i henhold til disse statistiske sjældne score.
DNA-ekstraktion og genotypebestemmelse
DNA blev ekstraheret fra 20 ml mundskylleprøver ved hjælp af Master Pure kit (Epicentre, Madison, vi). DRD4 48-bp VNTR var kendetegnet ved en PCR – amplifikationsprocedure med følgende primere: F5′ – CTT CCT ACC CTG CCC GCT CAT GCT GCT GCT CTA CTGG – 3′ og R5′ – ACC ACCACC GGC AGG ACC CTC ATG GCC TTG CGC TC-3. PCR-reaktioner blev udført under anvendelse af 5 liter-Masterblanding (Thermo scientific), 2 liter-primere (0,5 liter), 0,6 liter-Mg/Cl2 (2,5 mM), 0.4 liter DMSO 5% og 1 liter vand til i alt 9 liter Totalvolumen, og yderligere 1 liter genomisk DNA blev tilsat til blandingen. Alle PCR-reaktioner blev udført på en Biometra T1-Termocycler (Biometra, g Pristtingem, Tyskland). PCR-reaktionsbetingelsen var som følger: forvarmningstrin ved 94,0 liter C i 5 minutter, 34 denatureringscyklusser ved 94,0 liter C i 30 sekunder, reannealing ved 55 liter C i 30 sekunder og forlængelse ved 72 liter C i 90 sekunder. reaktionen fortsatte med at holde ved 72 liter C i 5 minutter. Reaktionsblandingen blev derefter elektroforeseret på en 3% agarosegel (AMRESCO) med ethidiumbromid for at screene for genotyper.fordelingen af genotypefrekvensen var i overensstemmelse med Hardy-Veinberg-ligevægten (chi-kvadrat = 0,01, p-værdi = 0,92).
statistisk analyse
Genotype blev klassificeret efter tilstedeværelsen eller fraværet af 7R-allelen af DRD4 (7 vs. no7) som i tråd med tidligere rapporter i litteraturen (ding et al., 2002). Gruppeforskelle på kreativitetsscore blev analyseret statistisk ved hjælp af ANOVAs med tilstedeværelsen (7) eller fravær (no7) af DRD4 7R-allelen som den uafhængige variabel. For at undersøge de forskellige aspekter af DT (flydende, fleksibilitet, originalitet) gennemførte vi en multivariat variansanalyse (MANOVA) separat for hver komponent.
resultater
for at bekræfte, at de to grupper (7, no7) ikke var signifikant forskellige med hensyn til alder og uddannelse, gennemførte vi uafhængige t-tests for at sammenligne de to grupper (Se tabel 1 For midler og standardafvigelser). Denne analyse viste, at de to grupper ikke var forskellige med hensyn til alder eller års uddannelse. Ikke-parametriske tests (Mann–Hvidney) afslørede signifikante forskelle i frekvensfordelingen for køn (å = 2,11, p < 0,05).
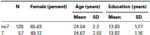
tabel 1. Alder, køn og uddannelsesdata for DRD4 7R-genotypen, SD: standardafvigelse.
de gennemsnitlige scoringer af de kreativitetsmål, der anvendes i de to opgaver, er opsummeret i tabel 2.

tabel 2. Midler og standardfejl (SE) for de to anvendte kreativitetsopgaver: AUT og Torrance.
flydende komponent
en multivariat ANOVA indikerede en generel effekt af DRD4 7R genotype for flydende komponent, F(2,182) = 3.25, p< 0.05. I betragtning af betydningen af den samlede test blev univariate hovedeffekter undersøgt. Signifikante univariate hovedeffekter for DRD4 genotype blev opnået for aut flydende F (1.183) = 4,95, p < 0.05(7R-allel, der udviser lavere flydende sammenlignet med no7-gruppen, figur 1A), men nåede ikke signifikansniveauet for Torrance-flydende F (1.183) = 2.95, p = 0.087.
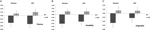
Figur 1. Kreativitet scorer på aut og Torrance opgaver adskilt til flydende (a), fleksibilitet (B) og originalitet (C). Midler blev omdannet til S-score.
Fleksibilitetskomponenten
multivariat ANOVA af fleksibilitetsmålene udviste en generel effekt for DRD4 7R genotype F(2.182) = 4.38, p < 0.05. Signifikante univariate hovedeffekter for DRD4-genotype blev opnået for aut-fleksibilitet F(1.183) = 4.57, p < 0.05 og for Torrance-fleksibilitet F (1.183) = 6.95, p < 0.01. Som vist i figur 1b udviste individer i 7R-allelgruppen i begge tilfælde lavere fleksibilitet sammenlignet med individer i no7-gruppen (tabel 2).
Originalitetskomponenten
multivariat ANOVA af originalitetsmålene udviste ingen signifikanseffekter, F(2.182) = 2.06, p = 0.13, skønt scorerne indikerede lavere originalitet for 7R-allelen i DRD4 (se figur 1C, tabel 2 for detaljer).
kønsforskelle
Diskussion
dt-opgaver kræver, at deltagerne leverer flere løsninger på et givet problem. I de opgaver, der blev brugt her, blev deltagerne forpligtet til at tænke på mange alternative mulige anvendelser til hverdagsgenstande (i AUT) og tegne flere tegninger med en bestemt form (cirkel) (i Torrance-opgaven). Resultaterne præsenteret her viser, at personer med DRD4 7R-allelen udviser lavere kreativitet målt ved dt. I betragtning af at 7R-allelen har været forbundet med dysfunktionel impulsivitet og dårlig hæmning (Congdon et al., 2008; Dreber et al., 2009), kan det faktum, at personer med 7R-allelen udviser lavere kreativitet, forklares ved deres manglende evne til at undertrykke eller hæmme åbenlyse (almindelige) svar, som angivet ved lavere originalitetsscore. Lavere fleksibilitet og flydende score kan indikere, at når et fælles svar er givet, bliver det sværere at skifte til en ny kategori (som angivet med de lave fleksibilitetsresultater), hvilket fører til Færre svar (som angivet med den lave flydende score). Yderligere bevis for forholdet mellem kreativitet og impulsivitet kan ses i sammenhængen mellem 7R-allelen og ADHD (Li et al., 2006) såvel som med øget impulsivitet hos ADHD-patienter (Langley et al., 2004). Interessant nok er kreativitet blevet knyttet til da-systemet i tilfælde af schisofreni (Eysenck, 1993). Selvom flere undersøgelser viser, at patienter med skisofreni og skisotypisk personlighed udviser høj kreativitet (O ‘ Reilly et al., 2001; Folley and Park, 2005), nyere modeller af schisofreni og kreativitet understøtter en omvendt U-formet forening, hvor kreativiteten kan være højere med lav-moderat schisotypisk egenskab, men falder, når sværhedsgraden af psykopatologi i schisofreni øges (Tsakanikos og Claridge, 2005; Stoneham og Coughtrey, 2009; Nelson og rav, 2010). Ja, i en nylig undersøgelse Jarac et al. (2012) rapporterede, at patienter med schisofreni viser nedsat kreativitet sammenlignet med kontroller. Desuden rapporterede forfatterne, at nedsat kreativitet var forbundet med lav kognitiv fleksibilitet og impulsivitet, afspejlet af deres lave score på Kortsorteringstesten. Disse resultater er i tråd med tidligere undersøgelser, der viser, at patienter med kronisk schisofreni udviser lavere kreativitet, som i tilfælde af kreativ flydende blev formidlet af scoringer på udøvende kontrolopgaver og i tilfælde af originalitet ikke var (Abraham et al., 2007). Kollektivt, mens mild funktionel impulsivitet kan forbedre kreativiteten blandt patienter med skisotypiske træk, især ved generering af ideer, der adskiller sig fra de givne eksempler (Abraham og Vindmann, 2008), kan den dysfunktionelle impulsivitet, der observeres ved svær skisofreni, dæmpe kreative evner.
resultaterne af den aktuelle undersøgelse understøtter forholdet mellem kreativitet og varianter af da genetiske polymorfier (Reuter et al., 2006; Runco et al., 2011). Desuden viser resultaterne i vores prøve, at DRD4-7R-allelens hovedvirkning på kreativitet kan skyldes dens indvirkning på fleksibilitet. Flere undersøgelser har forbundet kognitiv fleksibilitet med da-veje (Ashby og Isen, 1999; Dreisbach og Goschke, 2004; M., 2007). Det er blevet postuleret, at aktivering af D2-lignende receptorer (dvs.D2, D3 og D4) nedsætter Gabaergisk inhibering og letter aktivering af flere repræsentationer, hvilket øger fleksibiliteten som manifesteret ved sætskiftende evner (M., 2007; Seamans and Robbins, 2010). Desuden har D2-lignende receptorer været forbundet med fleksibel integration af ny information (2008). I tråd med dette, Reuter et al. (2006) fandt en sammenhæng mellem dopaminreceptoren D2 og kreativitet målt ved et sammensat indeks, selvom det ikke kan bestemmes, om fleksibilitet spillede en vigtig rolle i disse resultater. Derudover, Runco et al. (2011) rapporterede forskelle i kreativ flydende blandt bærere af flere dopamingener, blandt dem DRD4. Det fremgår imidlertid ikke af deres rapport, hvilken gruppe der udviser større kreative evner. Det er vigtigt at bemærke, at I modsætning til de resultater, der præsenteres her, Runco et al. (2011) fandt ikke en sammenhæng mellem mål for fleksibilitet målt ved dt-opgaver og DRD4. Denne inkonsekvens kan forklares ved forskelle i de anvendte populationer og gruppestørrelser eller ved forskelle i allelfrekvenser inden for den stikprøveudtagne population. Desuden er der tegn på, at sammenhængen mellem dopamin og DT muligvis ikke er lineær (Akbari Chermahini og Hommel, 2010, 2012), hvilket indikerer, at en interaktion mellem flere da-gener kan påvirke de individuelle resultater af hver genetisk variabilitet i sammenhæng med DT. Derudover har flere undersøgelser peget på det faktum, at da-veje kan være kønsfølsomme (Tammim L. R. og M. R. R. R., 2011; Vang et al., 2012). Jong et al. (2010) fandt for eksempel, at køn modulerede sammenhængen mellem DRD4 ekson3 polymorfisme og følelse af retfærdighed. I den aktuelle undersøgelse fandt vi ikke en interaktionseffekt mellem DRD4-7R og køn, hvilket indikerer, at køn ikke har en modulerende effekt på sammenhængen mellem DRD4 og kreativitet i vores prøve. Desuden indikerer resultaterne her en mangel på kønsforskelle i kreativitet og især i DT (til gennemgang, se Baer og Kaufman, 2008).
afslutningsvis foreslår vi i den aktuelle undersøgelse, at sammenhængen mellem DRD4 og kreativitet hovedsageligt er påvirket af fleksibilitet. Resultaterne af den aktuelle undersøgelse er i tråd med tidligere resultater, der peger på da-systemets involvering i kreativitet og tilføjer til den akkumulerende viden, der tyder på genetiske påvirkninger i kreativitet.
der er flere begrænsninger i undersøgelsen, der bør anerkendes. For det første undersøgte den aktuelle undersøgelse kun sammenhængen mellem en genetisk variabilitet i da—systemet—DRD4-og DT. Imidlertid, der er andre vigtige polymorfier i striatal DA-systemet, der kunne være forbundet med DT og kan bidrage både på en enkeltstående måde såvel som i gen-gen-interaktioner. En sådan polymorfisme er dopamintransportøren (DAT1), som har vist sig at være forbundet med inhibering af tilbagevenden., 2010) samt med impulsivitetsmålinger (2012). Som nævnt ovenfor er en anden mulighed, der ikke blev undersøgt her, de mulige gen-gen-interaktionseffekter på DT. Akbari Chermahini og Hommel (2010) rapporterede en ikke-lineær sammenhæng mellem en markør for striatal dopamin og DT, hvilket indikerer, at forholdet kan være mere komplekst, således at en polymorfisme af et gen kan påvirke DT gennem mægling af en anden genpolymorfisme. Fremtidige undersøgelser bør således overveje at undersøge interaktionen mellem disse to polymorfier i sammenhæng med fleksibilitet og DT.
interessekonflikt Erklæring
forfatterne erklærer, at forskningen blev udført i mangel af kommercielle eller økonomiske forhold, der kunne fortolkes som en potentiel interessekonflikt.
Akbari Chermahini, S., og Hommel, B. (2010). (B) forbindelsen mellem kreativitet og dopamin: spontane øjenblinkhastigheder forudsiger og adskiller divergerende og konvergerende tænkning. Kognition 115, 458-465. doi: 10.1016 / j. kognition.2010.03.007
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Cools, R. (2008). Dopamins rolle i den motiverende og kognitive kontrol af adfærd. Neuroscientist 14, 381-395. doi: 10.1177 / 1073858408317009
Pubmed abstrakt | Pubmed Fuld tekst | CrossRef Fuld tekst
Frank, M. J. (2005). Dynamisk dopaminmodulation i de basale ganglier: en neurocomputational redegørelse for kognitive underskud i medicinsk og ikke-medicinsk Parkinsonisme. J. Cogn. Neurosci. 17, 51–72. doi: 10.1162/0898929052880093
Pubmed abstrakt | Pubmed Fuld tekst/CrossRef Fuld tekst
Healey, D., and Rucklidge, J. J. (2006). En undersøgelse af forholdet mellem ADHD symptomatologi, kreativitet og neuropsykologisk funktion hos børn. Barn Neuropsykol. 12, 421–438. doi: 10.1080/09297040600806086
Pubmed abstrakt | Pubmed Fuld tekst/CrossRef Fuld tekst
Runco, M. A. og Richards, R. (1997). Fremtrædende kreativitet, dagligdags kreativitet og sundhed. Grøn, CT: Ableks.
Torrance, E. (1974). Forskelle er ikke underskud. Lære. Coll. Rec. 75, 471–487.