- introduktion
- materialer og metoder
- plantematerialer og væksttilstand
- PSI-og PSII-målinger
- Elektrokromisk Skiftanalyse
- Fotoinhibitoriske behandlinger
- statistisk analyse
- resultater
- Diskussion
- CEF-Stimuleringens rolle ved svagt lys
- ændring i P700-Iltningsforhold ved lavt lys er ikke relateret til pmf
- Etrii styrer P700 Iltningsforhold ved svagt lys
- konklusion
- Forfatterbidrag
- finansiering
- interessekonflikt Erklæring
introduktion
selvom lys er drivkraften for fotosyntese, kan højt lys forårsage betydelig fotoinhibition af photosystem II (PSII) for blade af skyggeetablerende planter (Kitao et al., 2000; Barth et al., 2001; Krause et al., 2004; Huang et al., 2015b, 2016b, c). Imidlertid genopretter fotoinhiberet PSII hurtigt ved svagt lys (He and Chov, 2003; Jang og Scheller, 2004, Allakhverdiev et al., 2005) på grund af den hurtige omsætningshastighed for D1-protein (Aro et al., 1993; Jang og Scheller, 2004; Allakhverdiev et al., 2005). Hastigheden af PSII-reparation blev reduceret ved inhibering af syntesen af ATP enten via PSI eller PSII (Allakhverdiev et al., 2005), hvilket indikerer, at den hurtige reparation af fotobeskadigede PSII-komplekser har brug for en stor mængde ATP på kort tid. En tidligere undersøgelse viste, at efter afkøling-induceret fotoinhibition af PSII, cyklisk elektronstrøm (CEF) blev signifikant stimuleret under genopretning ved lavt lys (Huang et al., 2010). Det blev foreslået, at denne stimulering af CEF ved svagt lys forbedrede syntesen af ATP til hurtig reparation af PSII (Huang et al., 2010). Imidlertid er der behov for flere beviser for at afklare den specifikke rolle CEF-stimulering ved svagt lys efter PSII-fotoinhibition.
under CEF cykles elektroner fra enten NADPH eller ferredoksin omkring PSI ind i plastokinonpuljen. Denne elektronoverførsel er koblet til protontranslokation og genererer en protongradient på tværs af thylakoidmembraner (Lyraph) (Shikanai og Yamamoto, 2017). Ud over at bidrage til ATP-syntese er en anden funktion af RPF nedregulering af fotosyntetisk elektrontransport ved forsuring af thylakoidlumen (Shikanai, 2014, 2016). Denne regulering involverer to forskellige mekanismer: den ene er knyttet til termisk energispredning og spreder overskydende absorberet lysenergi som varme fra PSII-antenner (Takahashi et al., 2009), og den anden er nedregulering af Cyt b6/f kompleks aktivitet og styrer hastigheden af elektronoverførsel til PSI (Suorsa et al., 2012, 2016; Tikkanen og Aro, 2014). I angiospermer inklusive Arabidopsis thaliana opererer to veje til PSI cyklisk elektrontransport (Shikanai, 2007). Den første CEF-vej er PGR5- / PGRL1-afhængig vej følsom over for antimycin a (Munekage et al., 2002; Sugimoto et al., 2013), og den anden er afhængig af chloroplast NADH dehydrogenase-lignende (NDH) kompleks (huler et al., 1998; Shikanai et al., 1998). Bidraget fra den pgr5- / PGRL1-afhængige vej er mere signifikant i C3-planter. Under højt lys inducerer aktiveringen af pgr5-/PGRL1-afhængig CEF forsuringen af thylakoidlumen og fører således til de høje niveauer af P700-iltningsforhold (Suorsa et al., 2012, 2016; Kono et al., 2014; Yamori et al., 2016). Til sammenligning var P700-iltningsforholdet ved svagt lys lidt påvirket af manglen i PGR5-/PGRL1-afhængig CEF (Munekage et al., 2002, 2004; Kono et al., 2014). Ved moderat PSII-fotoinhibition blev stimuleringen af CEF ved svagt lys ledsaget af en stigning i P700-iltningsrationen (Huang et al., 2010). Det er imidlertid uklart, om dette øgede P700-iltningsforhold skyldes CEF-stimuleringen.
ud over CEF spiller elektronoverførsel fra PSII (ETRII) en vigtig rolle i at påvirke tilstanden af PSI (Tikkanen et al., 2014; Huang et al., 2016a, c; Suorsa et al., 2016). I pgr5-planter af A. thaliana vil alvorlig fotoinhibition af PSII sandsynligvis fungere som den ultimative kontrol af fotosyntetisk elektronoverførsel, der tillader vedligeholdelse af P700 optimalt iltet under overskydende lys (Tikkanen et al., 2014). For nylig, Suorsa et al. (2016) viste, at pgr5-planter viste et højt P700-iltningsforhold under højt lys, da ETRII blev reduceret ved at kombinere knockout-mutationer i PsbO1, PsbP2, Psbk1, Psbk2 og PsbR loci. I afkølede tobaksblade deprimerede moderat PSII-fotoinhibition elektronstrømmen til PSI og øgede derefter P700-iltningsforholdet under yderligere kølebehandlinger (Huang et al., 2016a). Desuden førte chilling-induceret fotoinhibition af PSII til en depression af ETRII, som blev ledsaget af en stigning i P700-iltningsforholdet ved lavt lys (Huang et al., 2010). Vi antager, at P700-iltningsforholdet ved lavt lys primært styres af elektronstrøm fra PSII.
i vores tidligere rapport observerede vi, at den skyggeetablerende plante panaks notoginseng viste selektiv fotoinhibition af PSII under højlysspænding. Efter kortvarig eksponering for stress med højt lys faldt ETRII ved lavt lys markant, men CEF blev signifikant stimuleret. Samtidig steg P700-iltningsforholdet stort set. Vores specifikke mål var at (1) undersøge, om CEF-stimulering ved svagt lys hovedsageligt Letter syntesen af ATP; og (2) afgøre, om stigningen i P700-iltningsforholdet ved moderat PSII-fotoinhibition er mere relateret til ETRII snarere end CEF-stimuleringen eller ændringen i pmf. For at løse disse spørgsmål blev intakte blade af panaksa notoginseng behandlet ved 2258 lmol fotoner m-2 s-1 i 30 min. Før og efter behandling med højt lys blev energifordelingen i PSII, tilstanden af PSI og protonmotivkraft (PMF) ved et lavt lys på 54 liter-fotoner m-2 s-1 bestemt.
materialer og metoder
plantematerialer og væksttilstand
i denne undersøgelse blev 2-årige planter af en skyggeetablerende plante brugt til forsøg. Disse planter blev dyrket i let tilstand med 10% sollys (med maksimal lysintensitet midt på dagen på cirka 200 liter fotoner m-2 s-1). Der blev ikke oplevet vand-og ernæringsbelastninger for disse planter. 9 uger gamle fuldt udvidede blade blev brugt til fotosyntetiske målinger.
PSI-og PSII-målinger
fotosystem I-og PSII-parametre blev målt til 25 liter C ved samtidig registrering ved hjælp af et dobbelt Pam-100-målesystem. Klorofylfluorescensparametrene blev beregnet som følger: Fv/Fm = (Fm – Fo)/Fm, Y(II) = (Fm’ – Fs)/Fm’ (Genty et al., 1989), NPK = (Fm – Fm’)/Fm’. Fo og Fm er henholdsvis den mindste og maksimale fluorescens efter mørk akklimatisering. Fs er den lysindrettede steady-state fluorescens. Fo og Fm blev bestemt efter mørk akklimatisering i 30 minutter før og efter højlysbehandling. PSI fotosyntetiske parametre blev målt ved dobbelt PAM-100 baseret på P700 signal (forskel på intensiteter på 830 og 875 nm pulsmoduleret målelys, der nåede fotodetektoren) (Klughammer og Schreiber, 2008). P700 + signalerne (P) kan variere mellem et minimalt (P700 fuldt reduceret) og et maksimalt niveau (P700 fuldt iltet). Det maksimale niveau (Pm) blev bestemt ved anvendelse af en mætningsimpuls (600 ms og 10000 liter fotoner m-2 s-1) efter forbelysning med langt rødt lys, og Pm blev brugt til at estimere PSI-aktiviteten. Pm ‘ blev bestemt svarende til Pm, men med aktinisk lys i stedet for langt rødt lys. Kvanteudbyttet af PSI blev beregnet som Y (i) = (Pm’- P)/Pm. P700-iltningsforholdet i et givet aktinisk lys blev beregnet som Y (ND) = P/Pm. Kvanteudbyttet af PSI ikke-fotokemisk energidissipation på grund af acceptor-sidebegrænsning blev beregnet som Y (NA) = (Pm – Pm’)/Pm. Steady state-værdierne for Y (II), NPK, Y(I) og Y(ND) blev målt efter let akklimatisering ved svagt lys i 20 minutter.
Fotosyntetiske elektron flow gennem PSI og PSII blev beregnet som: ETRII = Y (II) ×PPFD ×0.84 ×0.5 (Krall og Edwards, 1992), ETRI = Y (I) ×PPFD×0.84 ×0.5 (Yamori et al., 2011), hvor 0,5 antages at være andelen af absorberet lys, der når PSI eller PSII, og 0,84 antages at være absorptansen (fraktionen af det indfaldende lys absorberet af blade). Omfanget af CEF-aktivering blev estimeret som etri/etrii-forhold (Yamori et al., 2011, 2015). Det skal bemærkes, at 1: 1-spændingspartitioneringen mellem PSI og PSII muligvis ikke er reel, fordi skyggeplanter har tendens til at øge PSII/PSI-forholdet (Lunde et al., 2003; Tikkanen et al., 2006; Grieco et al., 2012). Desuden blev PSI-og PSII-parametre i denne undersøgelse målt ved hjælp af rødt aktinisk lys (635 nm) leveret af Dual-PAM 100 (Vals, Tyskland). Fordi rødt aktinisk lys favoriserer ophidselse af PSII frem for PSI, kan spændingsenergifordelingen fra LHCII til PSII og PSI blive påvirket af det røde aktiniske lys (Tikkanen et al., 2017).
Elektrokromisk Skiftanalyse
ECS-signalet blev overvåget som absorbansændringen ved 515 nm ved hjælp af et Dual-PAM-100 (Vals, Effeltrich, Tyskland) udstyret med et p515 / 535 emitter-detektormodul (Vals). ECS-signalet blev opnået efter 20 min. belysning ved 54 prismol-fotoner m-2 s-1, bagefter blev ECS-henfaldet målt ved at slukke for det aktiniske lys i 30 s.analysen af ECS dark interval afslapningskinetik (DIRKECS) blev udført ved metoden fra Sacksteder et al. (2001) og et al. (2008). Total pmf blev estimeret ud fra den samlede amplitude af det hurtige henfald af ECS-signalet under 300 ms mørkt interval. Den langsomme afslapning af ECS-signal gjorde det muligt at genkende bidraget fra protongradient på tværs af thylakoidmembranerne (Prisph). Tidskonstanten for den første ordens ECS-afslapning (TEC ‘ er) er omvendt proportional med protonledningsevnen (gH+) af thylakoidmembranen gennem ATP-syntasen (Sacksteder og Kramer, 2000; , 2005). Som et resultat blev gH+ estimeret som den inverse af henfaldstidskonstanten .
Fotoinhibitoriske behandlinger
efter mørk akklimatisering i 30 minutter blev Fv / Fm og Pm målt i intakte blade. Bagefter blev disse intakte blade lysindrettet ved 59 liter-fotoner m-2 s-1 i 20 minutter, og parametre for klorofylfluorescens, P700-signal og ECS-signal blev registreret. Derefter blev det aktiniske lys ændret til 2258 lmol fotoner m-2 s-1. Efter eksponering for dette høje lys i 30 minutter blev det aktiniske lys straks ændret til 59 lysfotoner m-2 s-1, og de fotosyntetiske parametre blev registreret efter let akklimatisering i 20 minutter. Endelig blev Fv/Fm og Pm målt efter mørk akklimatisering i 30 minutter.
statistisk analyse
resultaterne blev vist som middelværdier for fem uafhængige eksperimenter. Uafhængig T-test blev anvendt ved signifikansniveau på LARP = 0,05 for at bestemme, om der var signifikante forskelle mellem forskellige behandlinger. Alle statistiske analyser blev udført ved hjælp af SPSS 16.0.
resultater
under fotosyntetisk induktion ved 59 liter-fotoner m-2 s-1 steg ETRII gradvist og nåede en stabil tilstand ved ca.18 min (figur 1a). Til sammenligning var etri konstant i denne induktionsfase (figur 1a). Efter begyndelsen af dette lave lys var etri meget højere end etrii. Efter en 20 minutters fotosyntetisk induktion var etri imidlertid lavere end etrii. Disse resultater foreslog aktivering af CEF i den indledende fase af induktion i overensstemmelse med tidligere undersøgelser (Joliot og Joliot, 2002, 2005; Makino et al., 2002). Efter denne fotosyntetiske induktion ved svagt lys blev bladene belyst ved et højt lys på 2258 liter fotoner m-2 s-1 i 30 minutter. Interessant nok faldt ETRII gradvist under højlysbehandlingen (figur 1b). I mellemtiden var værdien af etri højere end for ETRII (figur 1b), hvilket indikerer aktiveringen af CEF under højt lys. Efter denne højlysbehandling blev den maksimale foto-iltbare P700 (Pm) opretholdt stabil (figur 2a). Til sammenligning faldt den maksimale fluorescensintensitet (Fm) med 40%, og det maksimale kvanteudbytte af PSII (Fv/Fm) faldt fra 0,80 til 0,65 (figur 2b, C). Disse resultater indikerede den selektive fotoinhibition af PSII i blade af panaks notoginseng under højlysspænding.
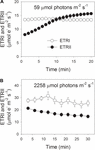
figur 1. Fotosyntetisk elektronstrøm ved lavt lys (A) og højt lys (B). (A) mørke akklimatiserede blade blev udsat for et svagt lys på 59 liter fotoner m-2 s-1 i 20 minutter. (B) efter belysning ved 59 liter-fotoner m-2 s-1 i 20 minutter for at aktivere elektronvasken i fotosyntese blev blade udsat for et højt lys på 2258 liter-fotoner m-2 s-1 i 30 minutter. Værdier er betyder, at se (n = 5).
før behandling med højt lys blev modne blade belyst ved et svagt lys på 59 liter-fotoner m-2 s-1 i 20 minutter for at aktivere fotosyntese, og derefter blev værdier for ETRI, ETRII, Y(ND) og NPK registreret. Efter højlysbehandling i 30 minutter blev værdier for ETRI, ETRII, Y(ND) og NPK registreret efter en ny akklimatiseringsfase på 20 minutter til svagt lys. Før behandling med højt lys var værdierne for etri og ETRII ved 59 liter-fotoner m-2 s-1 henholdsvis 13,4 og 15,7 liter-elektroner m-2 s-1 (figur 3a, B). Efter behandling med højt lys var ETRI og etrii ved svagt lys henholdsvis 12,8 og 11,2 liter-elektroner m-2 s-1 (figur 3a, B). Værdien af etrii ved 59 lp-fotoner m-2 s-1 faldt med ca.30% efter fotoinhibitorisk behandling, hvilket indikerer depression af ETRII ved moderat PSII-fotoinhibition. Før behandling med højt lys var værdien af etri / ETRII-forholdet ved 59 liter fotoner m-2 s-1 0,85 (figur 3c). Efter fotoinhibitorisk behandling steg etri / ETRII-forholdet signifikant til 1,14 (figur 3c). Disse resultater antydede stimulering af CEF ved svagt lys ved PSII-fotoinhibition (Yamori et al., 2011, 2015).
efter højlysbehandlingen ændrede steady-state-værdien af Y(I) ved 59 lp-fotoner m-2 s-1 ikke (figur 4a). Interessant nok faldt Y (NA) markant fra 0,36 til 0,23(figur 4b), og Y (ND) steg markant fra 0,1 til 0,25 (figur 4c). Disse resultater indikerede ændringen i PSI ‘ s tilstand ved svagt lys efter fotoinhibitorisk behandling. I mellemtiden ændrede værdien af NPK lidt (figur 4D). Som et resultat havde den moderate PSII-fotoinhibition forskellige effekter på P700-iltningsforhold og NPK ved svagt lys.
for at afklare, om stigningen i Y(ND) er forårsaget af en stigning i pmf, blev de elektrokromiske skiftsignaler ved 54 liter-fotoner m-2 s-1 bestemt efter let akklimatisering i 20 minutter før og efter højlysbehandlingen. Interessant nok faldt den samlede pmf signifikant med 18% efter højlysbehandling (figur 5A), men Pripph-niveauet ændrede sig ubetydeligt (figur 5b). Da dannelsen af pmf kan påvirkes af thylakoid protonledningsevne, blev protonledningsevnen (gH+) af thylakoidmembranen ved dette lave lys også undersøgt før og efter behandling med højt lys. Resultatet viste, at gH+ ikke ændrede sig med højlysbehandlingen (figur 5c), hvilket antyder, at højlysbehandlingen næppe påvirkede aktiviteten af chloroplast ATP-syntasen ved svagt lys. Fordi faldet på 30% i ETRII blev ledsaget af et fald på 18% i pmf, kompenserede stimuleringen af CEF ved svagt lys delvist for dannelsen af pmf og Rrph.

figur 5. Værdier for den samlede protonmotivkraft (pmf) (a), protongradienten (Priph) på tværs af thylakoidmembranerne (B) og protonledningsevnen (gH+) af thylakoidmembranen (C) ved svagt lys. Alle parametre blev målt efter tilpasning ved 54 liter fotoner m-2 s-1 i 20 minutter. Værdier er betyder, at se (n = 5). Asterisk angiver en signifikant (P < 0,05) ændring efter højlysbehandling.
Diskussion
CEF-Stimuleringens rolle ved svagt lys
det er blevet indikeret, at CEF spiller vigtige roller i opretholdelsen af fotosyntese og plantevækst (Yamori og Shikanai, 2016). Under højt lys menes CEF nu at være afgørende for at afbalancere ATP/NADPH-energibudgettet såvel som for at beskytte PSI og PSII mod skader (Munekage et al., 2002, 2004; Takahashi et al., 2009; Suorsa et al., 2012, 2016; et al., 2014; Huang et al., 2015a, 2017b). Ved lav lysintensitet spiller CEF en vigtig rolle i optimering af fotosyntetisk CO2-assimilering sandsynligvis via levering af ekstra ATP (Yamori et al., 2011, 2015; Nishikova et al., 2012; Huang et al., 2015a). CEF ‘ s hovedrolle moduleres fleksibelt som reaktion på lysintensitet. Især bidrager CEF hovedsageligt til at afbalancere ATP/NADPH-energibudgettet under submættende lysintensiteter, men beskytter hovedsageligt fotosyntetiske apparater mod fotoinhibition, når de udsættes for mættende lysintensiteter via forsuring af thylakoidlumen (Huang et al., 2015a). Interessant, Huang et al. (2010) fandt ud af, at CEF blev signifikant stimuleret ved svagt lys efter chilling-induceret fotoinhibition af PSII og antog, at denne CEF-stimulering hovedsageligt forbedrede syntesen af ATP til hurtig reparation af PSII. Der var imidlertid behov for flere beviser for at understøtte denne hypotese. I den foreliggende undersøgelse observerede vi, at værdien af etri/ETRII ved lavt lys steg signifikant (figur 3c), hvilket indikerer stimuleringen af CEF omkring PSI (Huang et al., 2011, 2012, 2017a, b; Yamori et al., 2011, 2015). Samtidig faldt amplituden af pmf signifikant, og Rrph faldt en smule. Disse resultater indikerede, at stimuleringen af CEF ikke udløste den RRF-afhængige nedregulering af fotosyntetisk elektrontransport. Med de andre ord letter denne stimulering af CEF ved svagt lys hovedsageligt syntesen af ATP.
selvom protonkonduktiviteten af chloroplast ATP-syntasen ikke blev påvirket af højlysbehandlingen, resulterede den mindre størrelse af pmf i et fald i den faktiske hastighed af ATP-syntese. Satserne for PSII-reparation og fotosyntetisk CO2-assimilering ved svagt lys er hovedsageligt begrænset af hastigheden af ATP-syntese (Allakhverdiev et al., 2005; Yamori et al., 2011, 2015; Nishikova et al., 2012). Fordi ETRII faldt signifikant ved PSII-fotoinhibition, blev hastigheden af ATP-syntese via etrii bemærkelsesværdigt reduceret. For at afbalancere ATP / NADPH-forholdet, der kræves af primær metabolisme og hurtig reparation af PSII, er der behov for en fleksibel mekanisme til at give ekstra ATP. Under denne betingelse faldt etrii med 30%, men den samlede pmf faldt med kun 18%. Disse resultater indikerer, at stimuleringen af CEF kompenserede for reduktionen i ETRII-afhængig dannelse af pmf, hvilket forbedrede syntesen af ATP. Den hurtige reparation af fotodamaged PSII er afhængig af ATP-syntese (Allakhverdiev et al., 2005). Acceleration af CEF omkring PSI øgede den intracellulære koncentration af ATP og accelererede således hastigheden af PSII-reparation i Synechocystis (Allakhverdiev et al., 2005). Samlet set foreslår vi, at stimulering af CEF ved svagt lys spiller en vigtig rolle i hurtig reparation af PSII-aktivitet via en yderligere ATP-syntese.
ændring i P700-Iltningsforhold ved lavt lys er ikke relateret til pmf
et interessant fænomen er, at P700-iltningsforholdet ved lavt lys steg signifikant ved moderat PSII-fotoinhibition (Huang et al., 2010), hvilket også er vist i den nuværende undersøgelse. Under forhold, hvor absorberet lys overstiger kravene til fotosyntese, regulerer CEF-afhængig generation af Kurph aktiviteten af Cyt b6/f kompleks aktivitet og styrer elektronstrømmen fra PSII til PSI (Suorsa et al., 2012, 2016; Shikanai, 2014, 2016), optimering af tilstanden af P700 i PSI og minimering af ROS-produktion i PSI under fotosyntese. I pgr5-planter af A. thaliana reduceres P700 med elektroner i lyset på grund af tabet af PMF-generation. I det pgr5-mutante akkumulerende flavodiironprotein blev niveauet af pmf imidlertid også gendannet til vildtypeniveauet, og P700 blev således iltet som i vildtypen (Yamamoto et al., 2016). I klorofyl B-mangelfulde hvedemutantlinjer fører utilstrækkelig thylakoid protongradient til overreduktion af PSI-acceptorsiden og dermed PSI-fotoinhibition under højt lys eller høj temperatur (Brestic et al., 2015, 2016). For nylig rapporterede nogle undersøgelser, at den øgede aktivitet af chloroplast ATP-syntasen nedsatte dannelsen af pmf og forårsagede overreduktion af fotosyntetisk elektrontransportkæde, hvilket resulterede i fotodamage af PSI under højt lys og svingende lys., 2017; Takagi et al., 2017). Disse rapporter understøtter PMF ‘ s kritiske rolle i at opretholde P700 optimalt iltet under overskydende lysenergi.
vores nuværende resultater viste, at efter højlysbehandlingen i 30 minutter faldt den samlede PMF,der blev dannet ved dette lave lys, signifikant, og dannelsen af Pripph ændrede sig lidt (figur 5A, B). I mellemtiden steg P700-iltningsforholdet signifikant fra 0,1 (før behandling) til 0,25 (efter behandling) (figur 4c). Disse resultater indikerer stærkt, at denne stigning i P700-iltningsforhold ikke kan forklares med ændringen i pmf, som stort set adskiller sig fra korrelationen mellem Y(ND) og pmf under højt lys (Yamamoto et al., 2016; Takagi et al., 2017). Derfor er reguleringseffekten af PMF på P700-iltningsforholdet minimal ved lavt lys, men bliver særlig afgørende under højt lys eller svingende lys. Under højt lys ledsages en høj værdi af Y(ND) normalt med en høj værdi af NPK (Munekage et al., 2002, 2004; Kono et al., 2014; TDC et al., 2015) på grund af stigningen i pmf. Nedskrivningen af dannelse af Kurpf fører til nedsatte niveauer af Y(ND) og NPK under højt lys (Suorsa et al., 2012; Kono et al., 2014; Yamamoto et al., 2016; et al., 2017). Induktionen af NPK ved svagt lys bestemmes hovedsageligt af niveauet af lumenforsuring. Efter behandling med højt lys ændrede omfanget af forsuring af lumen sig ikke (figur 5B), og NPK forblev stabil (figur 4D). Til sammenligning steg Y(ND) markant. Disse resultater indikerer, at ved svagt lys blev Y(ND) og NPK kontrolleret af forskellige reguleringsmekanismer.
Etrii styrer P700 Iltningsforhold ved svagt lys
PSI bliver kun overreduceret, når elektronstrømmen fra PSII overstiger PSI-elektronacceptorernes kapacitet til at klare elektronerne. Når elektronstrømmen til PSI er begrænset, er PSI ekstremt tolerant over for let stress i pgr5-planterne i A. thaliana (Suorsa et al., 2012, 2016; Tikkanen et al., 2014). For den skyggeetablerende plantepsykotria rubra var PSI-aktivitet ikke modtagelig for højlysspænding i nærvær af Dcmu (Huang et al., 2016c). I kølede tobaksblade tillod moderat PSII-fotoinhibition vedligeholdelse af P700 optimalt iltet og derefter beskyttet PSI-aktivitet mod yderligere fotodamage (Huang et al., 2016a). Disse resultater indikerede ETRII ‘s vigtige rolle i styringen af PSI’ s omkokstilstand under høje lys-eller kølelysspændinger. Vores nuværende resultater viste, at den moderate PSII-fotoinhibition førte til en signifikant depression af ETRII ved svagt lys. I mellemtiden ændrede PSI-aktiviteten og aktiviteten af chloroplast-ATP-syntasen sig lidt, og den samlede PMF og Kryph steg ikke. Som et resultat blev reduktionen i ETRII ikke forårsaget af den Kurfafhængige fotosyntetiske kontrol via Cyt b6/f-kompleks, men blev sandsynligvis induceret af faldet i PSII-aktivitet (Tikkanen et al., 2014). Den reducerede tilførsel af elektroner fra PSII til PSI førte til det øgede niveau af P700-iltning. Således blev P700-iltningsforholdet ved lavt lys stort set kontrolleret af ETRII.
konklusion
Vi fandt ud af, at den selektive fotoinhibition af PSII inducerede en stimulering af CEF og en stigning i P700-iltningsforholdet ved lavt lys. Stimuleringen af CEF udløste ikke den RRF-afhængige nedregulering af fotosyntetisk elektrontransport. Som et resultat letter denne stimulering af CEF ved svagt lys hovedsageligt syntesen af ATP, hvilket er afgørende for hurtig reparation af fotodamaged PSII. Stigningen i P700-iltningsforholdet kunne ikke forklares med ændringen i den Lutphafhængige fotosyntetiske kontrol ved Cyt B6/f-komplekset, men skyldtes primært den nedsatte tilførsel af elektroner fra PSII til PSI.
Forfatterbidrag
vi har udtænkt og designet forskningen. HH og Y-JY udførte eksperimenterne. HH, Y-JY, S-BS og TL analyserede dataene. H skrev det første udkast til manuskriptet, som blev intensivt redigeret af alle forfatterne.
finansiering
denne undersøgelse blev støttet af National Natural Science Foundation of China (31670343), Youth Innovation Promotion Association of the Chinese Academy of Sciences (2016347) og de store videnskabs-og Teknikprogrammer i Yunnan-provinsen (2016sf001).
interessekonflikt Erklæring
forfatterne erklærer, at forskningen blev udført i mangel af kommercielle eller økonomiske forhold, der kunne fortolkes som en potentiel interessekonflikt.
Kono, M., Noguchi, K. og Terashima, I. (2014). Roller af den cykliske elektronstrøm omkring PSI (CEF-PSI) og O2-afhængige alternative veje til regulering af den fotosyntetiske elektronstrøm i kortvarigt svingende lys i Arabidopsis thaliana. Plantecelle Physiol. 55, 990–1004. doi: 10.1093 / pcp / pcu033
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Shikanai, T. (2016). Regulatorisk netværk af protonmotivkraft: bidrag fra cyklisk elektrontransport omkring fotosystem I. Photosynth. Res. 129, 253-260. doi: 10.1007 / s11120-016-0227-0
PubMed abstrakt / CrossRef Fuld tekst/Google Scholar
Yamori, Makino, A. og Shikanai, T. (2016). En fysiologisk rolle cyklisk elektrontransport omkring fotosystem I ved opretholdelse af fotosyntese under svingende lys i ris. Sci. Rep. 6: 20147. doi: 10.1038/srep20147
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Yamori, V., og Shikanai, T. (2016). Fysiologiske funktioner ved cyklisk elektrontransport omkring fotosystem I til opretholdelse af fotosyntese og plantevækst. Annu. Rev. Plant Biol. 67, 81–106. doi: 10.1146 / annurev-arplant-043015-112002
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
Yamori, H., Shikanai, T. og Makino, A. (2015). Fotosystem I cyklisk elektronstrøm via chloroplast NADH dehydrogenase-lignende kompleks udfører en fysiologisk rolle for fotosyntese ved svagt lys. Sci. Rep. 5: 13908. doi: 10.1038/srep13908
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Jang, S. P. og Scheller, H. V. (2004). Fotoinhibition af fotosystem I ved køletemperatur og efterfølgende genopretning i Arabidopsis thaliana. Plantecelle Physiol. 45, 1595–1602. doi: 10.1093 / pcp / pch180
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar