- Introducción
- Método
- Participantes
- Evaluación de la creatividad
- Prueba de Pensamiento Creativo de Torrance (Subescala de Círculos)
- Tarea de usos alternativos (AUT)
- Prueba previa
- Extracción de ADN y genotipado
- Análisis estadístico
- Resultados
- El Componente de fluidez
- El Componente de flexibilidad
- El Componente de Originalidad
- Diferencias de género
- Discusión
- Declaración de Conflicto de Intereses
Introducción
La cognición creativa juega un papel importante en las artes, en la invención y la innovación, así como en la vida cotidiana (Runco y Richards, 1997). Aunque la creatividad se ha considerado una capacidad humana única, los comportamientos creativos espontáneos (p. ej., creando nuevas herramientas entre primates y aves) se ha demostrado que también ocurren en no humanos (Byrne y Bates, 2007), lo que atestigua aún más las profundas raíces evolutivas y biológicas de la creatividad. La creatividad se ha definido como la capacidad de producir respuestas novedosas (es decir, originales, raras e inesperadas) y adecuadas (es decir, adaptables y útiles de acuerdo con las limitaciones de la tarea) (Sternberg y Lubart, 1999). Uno de los enfoques psicométricos para medir la creatividad implica tareas de pensamiento divergente (DT) en las que se pide a los participantes que respondan a un problema determinado con múltiples soluciones (Dietrich y Kanso, 2010). Las pruebas de TD generalmente miden varios aspectos de la creatividad, incluida la fluidez creativa, la flexibilidad y la originalidad (Torrance, 1974). Por lo tanto, aunque no es sinónimo de creatividad, las tareas de TD proporcionan mediciones estructuradas y objetivas de la creatividad (Sternberg y Lubart, 1999; Jung et al., 2009). En particular, se ha demostrado que las puntuaciones en las tareas de TD están correlacionadas positivamente con medidas ecológicamente válidas de logro creativo (Carson et al., 2005), así como con creatividad autoevaluada (Furnham y Bachtiar, 2008).
Estudios previos han señalado la implicación del sistema dopaminérgico (DA)en la creatividad (Heilman et al., 2003; Flaherty, 2005; Takeuchi et al., 2010). Takeuchi et al. (2010) encontraron que las diferencias individuales en la creatividad, medidas por DT, se correlacionaban positivamente con la materia gris en las regiones del sistema DA, incluida la corteza prefrontal dorsolateral, los ganglios basales bilaterales, la sustancia negra y el área ventral tegmental. Además, varios estudios genéticos han demostrado una relación entre la DT y la neurotransmisión de dopamina (Reuter et al., 2006; De Manzano et al., 2010; Runco et al., 2011). Por ejemplo, Reuter et al. (2006) encontraron que la creatividad, medida por las tareas de DT que involucran creatividad tanto figurativa como verbal, está significativamente asociada con polimorfismos del gen receptor de dopamina D2 (DRD2). Además, varios estudios han encontrado que la dopamina cortical está involucrada en la flexibilidad cognitiva (Frank, 2005; Cools, 2008; Garcia-Garcia et al., 2010), uno de los principales componentes de DT.
Un gen candidato a DA interesante para la creatividad es el gen receptor de dopamina D4 (DRD4). El receptor DRD4 es uno de los cinco receptores de dopamina y desempeña un papel importante en la mediación de la señalización sináptica de dopamina. El gen se caracteriza por un número variable de pares de bases de 48 repeticiones en tándem (48-bp VNTR) ubicadas en la región codificante del tercer exón. DRD4 El polimorfismo VNTR de 48 pb varía de 2 a 11 repeticiones entre individuos (Asghari et al., 1995; Cravchik y Goldman, 2000), donde la 4-repetición (4R) es la repetición más común en poblaciones caucásicas y la 7-repetición (7R) es la segunda variante más común (Chang et al., 1996; Ding et al., 2002). Curiosamente, la 7R se ha asociado previamente con comportamientos creativos de la vida real, como el rasgo de personalidad que busca novedades (Ebstein et al., 1996). De hecho, se cree que la búsqueda de novedades-la tendencia hacia la actividad exploratoria-es una de las características de las personas creativas (Chavez-Eakle et al., 2006; Drago et al., 2009). Aunque la evidencia de una asociación entre DRD4 7R y la búsqueda de novedades es inconsistente, como lo demuestran varios informes de metanálisis (Kluger et al., 2002; Munafò et al., 2008), muchos estudios han encontrado una asociación significativa entre la búsqueda de novedades y el alelo 7R del DRD4 (Ebstein et al., 1996; Benjamin et al., 2000; Keltikangas-Järvinen et al., 2002; Becker et al., 2005). Roussos et al. (2009) han sugerido recientemente que las diferencias en la medición de la búsqueda de novedad utilizando escalas de autoinforme pueden explicar los resultados inconsistentes entre el polimorfismo 7R y la búsqueda de novedad. En línea con esto, se ha encontrado repetidamente que el sistema de DA juega un papel importante en el rasgo de personalidad de la búsqueda de novedades (Flaherty, 2005; Schweizer, 2006), lo que atestigua aún más el papel potencial de DRD4 en la creatividad.
Sin embargo, en contraste con el papel de DRD4 48-bp VNTR 7R en la búsqueda de novedades, la evidencia reciente indica que la 7R en realidad está asociada con un deterioro de la flexibilidad cognitiva, uno de los principales aspectos de la creatividad (Strobel et al., 2004; Congdon et al., 2008). La flexibilidad implica la capacidad de hacer una respuesta alternativa después de inhibir con éxito una respuesta actual. Los hallazgos hasta ahora sobre la asociación entre inhibición y DRD4 son mixtos. Congdon et al. (2008) encontraron que los participantes con el alelo 7R del DRD4 exhiben un mayor tiempo de reacción de la señal de parada (SSRT) en una tarea activa/nula, lo que refleja un control inhibitorio más pobre, mientras que Forbes et al. (2009) no encontraron tal efecto en la Escala de Impulsividad de Barratt (BIS), una herramienta de autoinforme para medir la impulsividad. Por otro lado, Colzato et al. (2010) demostraron que estos efectos mixtos pueden deberse al hecho de que estudios anteriores abordaron la impulsividad como un proceso monolítico, mientras que la impulsividad en realidad puede dividirse en tipos funcionales y disfuncionales de acuerdo con el Inventario de Impulsividad de Dickman (DII; Dickman, 1990). La impulsividad disfuncional es la tendencia a actuar sin previsión en los casos en que dicha acción es inapropiada, mientras que la impulsividad funcional es una tendencia similar implementada en situaciones apropiadas (Colzato et al., 2010). Colzato et al. (2010) encontraron que los individuos con DRD4 7R, que se asocia con niveles más altos de DA estriado, exhibieron una impulsividad disfuncional más alta. Varios estudios meta-analíticos han señalado al alelo 7R del DRD4 como un alelo de riesgo para el trastorno por déficit de atención con hiperactividad (TDAH; Faraone et al., 2001; DiMaio et al., 2003), un trastorno caracterizado por una impulsividad disfuncional elevada (Young et al., 2007). Además, la relación entre el TDAH y la creatividad ha sido investigada en varios estudios (Shaw y Brown, 1990; Healey y Rucklidge, 2006), aunque hasta ahora estos estudios han arrojado resultados poco claros. Mientras que algunos han encontrado que el TDAH y su sintomatología están correlacionados positivamente con la creatividad (Healey y Rucklidge, 2006; White y Shah, 2011), otros han reportado una tendencia opuesta (Funk et al., 1993; Healey y Rucklidge, 2005, 2008). Curiosamente, el TDA / H está asociado en el comportamiento con deficiencias en el control ejecutivo, incluido el cambio de configuración de atención (Boonstra et al., 2005), y flexibilidad (Barkley et al., 1997; Sergeant et al., 2002), ambos importantes para la flexibilidad cognitiva, una dimensión central de la creatividad (Dietrich, 2004; Durstewitz y Seamans, 2008). Estos hallazgos indican que los individuos con el alelo 7R de la DRD4 pueden exhibir niveles más bajos de creatividad y flexibilidad disminuida en las tareas de DT en particular. Por lo tanto, el exón III DRD4 VNTR es un candidato biológicamente plausible para contribuir a las diferencias individuales en la creatividad.
En conjunto, parece que, por un lado, el alelo 7R, como alelo de riesgo para las funciones ejecutivas deterioradas, la flexibilidad y el cambio de configuración, puede estar asociado con una baja creatividad. Por otro lado, la asociación entre el alelo 7R y la búsqueda de novedad superior indica que los individuos con el alelo 7R pueden exhibir una mayor flexibilidad y creatividad.
El presente estudio fue diseñado para explorar el complejo papel del DRD4 7R en la configuración parcial de la creatividad humana. Para caracterizar mejor esta relación, evaluamos dos tipos de pruebas de TD (figurativa y verbal). La puntuación de cada tarea se dividió de acuerdo con las tres dimensiones principales de DT, a saber, fluidez creativa, flexibilidad y originalidad, y cada dimensión se analizó por separado para probar la hipótesis de que DRD4 afecta diferencialmente a cada dimensión de la creatividad.
Método
Participantes
La muestra estuvo compuesta por 185 estudiantes de la Universidad de Haifa en Israel (112 mujeres). Todos los participantes eran caucásicos (autoinformados), con una edad media de 24,5 años (DE ± 2,1). Los participantes fueron reclutados a través de anuncios publicados en la universidad. Todos los participantes eran voluntarios remunerados.
Evaluación de la creatividad
La creatividad se evaluó mediante dos pruebas DT, una subescala de la subprueba figurativa de la prueba de pensamiento creativo de Torrance (Torrance, 1974) y la Tarea de Usos Alternativos (AUT; Guilford et al., 1978). Ambas pruebas incluyeron las tres dimensiones principales de DT, a saber, flexibilidad, originalidad y fluidez.
Prueba de Pensamiento Creativo de Torrance (Subescala de Círculos)
A los participantes se les dio una página en la que se dibujaron 30 círculos idénticos. Se les pidió que dibujaran tantos objetos significativos diferentes como fuera posible dentro de un límite de tiempo de 10 minutos, donde cada dibujo debe incluir al menos un círculo. La puntuación incluyó fluidez (número de respuestas producidas), flexibilidad (número de categorías) y originalidad, calculadas de acuerdo con la puntuación de las respuestas originales, como se detalla en la guía de puntuación de Pruebas de Pensamiento Creativo de Torrance (Torrance, 1974).
Tarea de usos alternativos (AUT)
A los participantes se les dio una lista de cinco objetos comunes y se les pidió que enumeraran tantos usos alternativos como fuera posible para cada objeto, dentro de un límite de tiempo de 10 minutos. El uso diario más común se indicó entre paréntesis. Los objetos eran: zapato( uso común: desgaste a pie); botón (uso común: cierre de objetos); lápiz (uso común: dibujo o escritura); neumático (uso común: rueda de automóvil); y vaso para beber (uso común: contiene líquido). Solo se contaron e incluyeron las respuestas que no reiteraban los usos comunes dados. Al igual que en la prueba de Torrance, la puntuación incluía fluidez, flexibilidad y originalidad. Dado que no hay pautas para la puntuación de las respuestas originales en el AUT, las respuestas originales se definieron como respuestas estadísticamente infrecuentes de acuerdo con una prueba previa realizada en nuestro laboratorio.
Prueba previa
Con el fin de crear un criterio válido de frecuencia de respuesta, un grupo de 100 participantes sanos que no participaron en este estudio completaron el AUT. Para cada objeto, se recopiló una lista de todos los usos posibles de todos los participantes. A partir de esta lista se calculó una medida estadística de infrecuencias para evaluar la puntuación de originalidad de cada respuesta y, posteriormente, de cada participante. A las respuestas se les asignó una puntuación de cero si el 5% o más de los participantes enumeraban un uso dado, una puntuación de uno si entre el 2% y el 4,99% de los participantes lo enumeraban, y una puntuación de dos si menos del 1,99% enumeraba el uso. Se calculó un puntaje promedio de originalidad para cada participante de acuerdo con estos puntajes estadísticos de infrecuencia.
Extracción de ADN y genotipado
El ADN se extrajo de 20 ml de muestras de enjuague bucal utilizando el Master Pure kit (Epicentre, Madison, WI). El DRD4 VNTR de 48 pb se caracterizó por un procedimiento de amplificación de PCR con los siguientes cebadores: F5 ‘- CTT CCT ACC CTG CCC GCT CAT GCT GCTGCT CTA CTGG – 3’ y R5 ‘ – ACC ACCACC GGC AGG ACC CTC ATG GCC TTG CGC TC – 3. Las reacciones de PCR se llevaron a cabo utilizando Mezcla Maestra de 5 µl (Thermo scientific), cebadores de 2 µl (0,5 µM), 0,6 µl Mg/Cl2 (2,5 mm), 0.se añadieron a la mezcla 4 µl de DMSO al 5% y 1 µl de agua hasta un volumen total de 9 µl, y 1 µl adicional de ADN genómico. Todas las reacciones de PCR se llevaron a cabo en un termociclador Biometra T1 (Biometra, Güttingem, Alemania). La reacción de PCR condición fue la siguiente: precalentamiento paso en 94.0°C por 5 min, 34 ciclos de desnaturalización a 94.0°C por 30 s, reannealing a 55°C por 30 s y extensión a 72°C durante 90 seg. La reacción se procedió a realizar a 72°C por 5 min. La mezcla de reacción se electroforizó en un gel de agarosa al 3% (AMRESCO) con bromuro de etidio para detectar genotipos.
La distribución de la frecuencia del genotipo fue de acuerdo con el equilibrio de Hardy-Weinberg (chi-cuadrado = 0,01, valor de p = 0,92).
Análisis estadístico
El genotipo se clasificó de acuerdo con la presencia o ausencia del alelo 7R del DRD4 (7 vs.no7), de acuerdo con informes previos en la literatura (Ding et al., 2002). Las diferencias de grupo en las puntuaciones de creatividad se analizaron estadísticamente utilizando ANOVAs, con la presencia (7) o ausencia (no7) del alelo DRD4 7R como variable independiente. Para examinar los diferentes aspectos de la DT (fluidez, flexibilidad, originalidad), realizamos un análisis multivariado de varianza (MANOVA) por separado para cada componente.
Resultados
Para confirmar que los dos grupos (7, no7) no eran significativamente diferentes en términos de edad y educación, realizamos pruebas t independientes para comparar los dos grupos (ver Tabla 1 para medias y desviaciones estándar). Este análisis reveló que los dos grupos no diferían en cuanto a edad o años de educación. Las pruebas no paramétricas (Mann–Whitney) revelaron diferencias significativas en la distribución de frecuencias por sexo (Z = 2,11, p < 0,05).
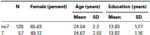
la Tabla 1. Datos de edad, sexo y educación para el genotipo DRD4 7R, DE: desviación estándar.
Las puntuaciones medias de las medidas de creatividad utilizadas en las dos tareas se resumen en la Tabla 2.

la Tabla 2. Medios y Error estándar (SE) de las dos tareas de creatividad utilizadas: AUT y Torrance.
El Componente de fluidez
Un ANOVA multivariado indicó un efecto general del genotipo DRD4 7R para el componente de fluidez, F(2,182)=3.25, p < 0.05. Dada la importancia de la prueba global, se examinaron los principales efectos univariados. Se obtuvieron efectos principales univariados significativos para el genotipo DRD4 para la fluidez AUT F(1183) = 4,95, p < 0.05(alelo 7R con menor fluidez en comparación con el grupo no7, Figura 1A), pero no alcanzó el nivel de significación para la fluidez de Torrance F (1.183) = 2,95, p = 0,087.
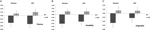
la Figura 1. Las puntuaciones de creatividad en las tareas AUT y Torrance se dividen en fluidez (A), flexibilidad (B) y originalidad (C). Los medios se transformaron en puntuaciones Z.
El Componente de flexibilidad
ANOVA multivariante de las medidas de flexibilidad mostró un efecto general para el genotipo DRD4 7R F(2.182) = 4.38, p < 0,05. Importante univariado de efectos principales para DRD4 genotipo fueron obtenidos para AUT flexibilidad F(1,183) = 4.57, p < 0,05 y para Torrance flexibilidad F(1,183) = 6.95, p < 0.01. Como se muestra en la Figura 1B, en ambos casos los individuos del grupo alelo 7R mostraron menor flexibilidad en comparación con los individuos del grupo no7 (Tabla 2).
El Componente de Originalidad
ANOVA Multivariante de las medidas de originalidad no mostró ningún efecto de significación, F(2.182) = 2,06, p = 0.13, aunque las puntuaciones indicaron menor originalidad para el alelo 7R de la DRD4 (ver Figura 1C, Tabla 2 para más detalles).
Diferencias de género
Discusión
Las tareas de DT requieren que los participantes proporcionen múltiples soluciones a un problema determinado. En las tareas utilizadas aquí, se pidió a los participantes que pensaran en muchos usos alternativos posibles para los artículos cotidianos (en el AUT) y que dibujaran varios dibujos que incorporaran una forma específica (círculo) (en la tarea Torrance). Los resultados presentados aquí demuestran que los individuos con el alelo DRD4 7R exhiben menor creatividad medida por DT. Considerando que el alelo 7R se ha asociado con impulsividad disfuncional e inhibición deficiente (Congdon et al., 2008; Dreber et al., 2009), el hecho de que los individuos con el alelo 7R exhiban menor creatividad puede explicarse por su incapacidad para suprimir o inhibir respuestas obvias (comunes), como lo indican las puntuaciones de originalidad más bajas. Los puntajes de flexibilidad y fluidez más bajos pueden indicar que una vez que se ha dado una respuesta común, se hace más difícil cambiar a una nueva categoría (como se indica en los puntajes de flexibilidad bajos), lo que lleva a menos respuestas (como se indica en el puntaje de fluidez bajo). Otra evidencia de la relación entre creatividad e impulsividad se puede ver en la asociación del alelo 7R con el TDAH(Li et al., 2006), así como con mayor impulsividad en pacientes con TDAH (Langley et al., 2004). Curiosamente, la creatividad se ha vinculado al sistema de DA en el caso de la esquizofrenia (Eysenck, 1993). Aunque varios estudios muestran que los pacientes con esquizofrenia y personalidad esquizotípica exhiben una alta creatividad(O’Reilly et al., 2001; Folley y Park, 2005), los modelos recientes de esquizofrenia y creatividad apoyan una asociación en forma de U invertida en la que la creatividad puede ser mayor con el rasgo esquizotípico bajo-moderado, pero disminuye a medida que aumenta la gravedad de la psicopatología en la esquizofrenia (Tsakanikos y Claridge, 2005; Stoneham y Coughtrey, 2009; Nelson y Rawlings, 2010). De hecho, en un estudio reciente Jaracz et al. (2012) informaron que los pacientes con esquizofrenia muestran una menor creatividad en comparación con los controles. Además, los autores informaron que el deterioro de la creatividad se asoció con una baja flexibilidad cognitiva e impulsividad, lo que se refleja en sus bajas puntuaciones en la Prueba de Clasificación de tarjetas de Wisconsin (WCST). Estos resultados están en línea con investigaciones anteriores que muestran que los pacientes con esquizofrenia crónica exhiben menor creatividad, que en el caso de la fluidez creativa estaba mediada por puntuaciones en tareas de control ejecutivo y en el caso de la originalidad no (Abraham et al., 2007). Colectivamente, mientras que la impulsividad funcional leve puede mejorar la creatividad entre los pacientes con rasgos esquizotípicos, especialmente al generar ideas que difieren de los ejemplos dados (Abraham y Windmann, 2008), la impulsividad disfuncional observada en la esquizofrenia grave puede disminuir las habilidades creativas.
Los resultados del estudio actual apoyan la relación entre la creatividad y las variantes de polimorfismos genéticos DA (Reuter et al., 2006; Runco et al., 2011). Además, los resultados encontrados en nuestra muestra indican que el principal efecto del alelo DRD4-7R en la creatividad puede deberse a su impacto en la flexibilidad. Varios estudios han relacionado la flexibilidad cognitiva con las vías de DA (Ashby e Isen, 1999; Dreisbach y Goschke, 2004; Müller et al., 2007). Se ha postulado que la activación de receptores similares a D2 (es decir, D2, D3 y D4) disminuye la inhibición gabaérgica y facilita la activación de múltiples representaciones, aumentando así la flexibilidad manifestada por las habilidades de cambio de configuración (Müller et al., 2007; Seamans y Robbins, 2010). Además, los receptores similares a D2 se han vinculado a la integración flexible de nueva información (Durstewitz y Seamans, 2008). En línea con esto, Reuter et al. (2006) encontraron una asociación entre el receptor de dopamina D2 y la creatividad medida por un índice compuesto, aunque no se puede determinar si la flexibilidad jugó un papel importante en estos resultados. Además, Runco et al. (2011) reportaron diferencias en la fluidez creativa entre portadores de varios genes dopaminérgicos, entre ellos el DRD4. Sin embargo, no está claro en su informe qué grupo exhibe mayores habilidades creativas. Es importante señalar que, a diferencia de los resultados presentados aquí, Runco et al. (2011) no encontraron una asociación entre las medidas de flexibilidad medidas por las tareas de DT y DRD4. Esta inconsistencia puede explicarse por diferencias en las poblaciones utilizadas y el tamaño de los grupos o por diferencias en las frecuencias de los alelos dentro de la población muestreada. Además, hay evidencia que sugiere que la asociación entre dopamina y DT puede no ser lineal (Akbari Chermahini y Hommel, 2010, 2012), lo que indica que una interacción entre varios genes DA puede influir en los resultados individuales de cada variabilidad genética en el contexto de DT. Además, varios estudios han señalado el hecho de que las rutas de DA pueden ser sensibles al género (Tammimäki y Männistö, 2011; Wang et al., 2012). Zhong et al. (2010), por ejemplo, encontraron que el género modulaba la asociación entre el polimorfismo del exono3 DRD4 y el sentido de justicia. En el presente estudio no encontramos un efecto de interacción entre el DRD4-7R y el género, lo que indica que el género no tiene un efecto modulador en la asociación entre el DRD4 y la creatividad en nuestra muestra. Además, los resultados presentados aquí indican una falta de diferencias de género en la creatividad, y en particular en la TD (para una revisión, véase Baer y Kaufman, 2008).
En conclusión, en el presente estudio sugerimos que la asociación encontrada entre DRD4 y creatividad está influenciada principalmente por la flexibilidad. Los resultados del estudio actual están en línea con los resultados anteriores que apuntan a la participación del sistema de DA en la creatividad y se suman al cuerpo acumulado de conocimiento que sugiere influencias genéticas en la creatividad.
Hay varias limitaciones del estudio que deben ser reconocidas. En primer lugar, el estudio actual solo examinó la asociación entre una variabilidad genética en el sistema de DA—el DRD4—y el DT. Sin embargo, hay otros polimorfismos importantes en el sistema DA estriado que podrían estar asociados con la TD y pueden contribuir tanto de manera independiente como en interacciones gene-gen. Uno de estos polimorfismos es el transportador de dopamina (DAT1), que ha demostrado estar asociado con la inhibición del retorno (Colzato et al., 2010), así como con medidas de impulsividad (Gizer y Waldman, 2012). Además, como se mencionó anteriormente, otra posibilidad que no se exploró aquí son los posibles efectos de interacción gene-gen en la TD. Akbari Chermahini y Hommel (2010) informaron una asociación no lineal entre un marcador de dopamina estriada y DT, lo que indica que la relación puede ser más compleja, de modo que un polimorfismo de un gen puede afectar DT a través de la mediación de otro polimorfismo génico. Por lo tanto, estudios futuros deberían considerar examinar la interacción entre estos dos polimorfismos en el contexto de la flexibilidad y la TD.
Declaración de Conflicto de Intereses
Los autores declaran que la investigación se realizó en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Akbari Chermahini, S., and Hommel, B. (2010). El (b) vínculo entre la creatividad y la dopamina: las tasas de parpadeo espontáneo predicen y disocian el pensamiento divergente y convergente. Cognición 115, 458-465. doi: 10.1016 / j. cognición.2010.03.007
Resumen de Pubmed / Texto Completo de Pubmed / Texto completo cruzado
Cools, R. (2008). Papel de la dopamina en el control motivacional y cognitivo del comportamiento. Neurocientífico 14, 381-395. doi: 10.1177/1073858408317009
Resumen de Pubmed | Texto Completo de Pubmed | Texto completo cruzado
Frank, M. J. (2005). Modulación dinámica de dopamina en los ganglios basales: una cuenta neurocomputacional de déficits cognitivos en parkinsonismo medicado y no medicado. J. Cogn. Neurociencia. 17, 51–72. doi: 10.1162 / 0898929052880093
Resumen de Pubmed / Texto completo de Pubmed / Texto completo cruzado
Healey, D., and Rucklidge, J. J. (2006). Una investigación sobre la relación entre la sintomatología del TDAH, la creatividad y el funcionamiento neuropsicológico en niños. Neuropsicol Infantil. 12, 421–438. doi: 10.1080 / 09297040600806086
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Runco, M. A., and Richards, R. (1997). Creatividad Eminente, Creatividad Cotidiana y Salud. Greenwich, CT: Ablex.
Torrance, E. (1974). Las diferencias no son déficits. Enseñar. Coll. Rec. 75, 471–487.