- Introduction
- Méthode
- Participants
- Évaluation de la créativité
- Test de Torrance de la Pensée créative (Sous-échelle des cercles)
- Tâche d’utilisations alternatives (AUT)
- Pré-test
- Extraction et génotypage de l’ADN
- L’analyse statistique
- Résultats
- La composante de fluidité
- La composante de flexibilité
- La Composante Originalité
- Différences entre les sexes
- Discussion
- Déclaration sur les conflits d’intérêts
Introduction
La cognition créative joue un rôle important dans les arts, dans l’invention et l’innovation, ainsi que dans la vie quotidienne (Runco et Richards, 1997). Bien que la créativité ait été considérée comme une capacité humaine unique, les comportements spontanément créatifs (p. ex., créant de nouveaux outils chez les primates et les oiseaux) se sont également produits chez des non-humains (Byrne et Bates, 2007), attestant davantage des racines évolutives et biologiques profondes de la créativité. La créativité a été définie comme la capacité de produire des réponses à la fois nouvelles (c’est-à-dire originales, rares et inattendues) et appropriées (c’est-à-dire adaptatives et utiles en fonction des contraintes de la tâche) (Sternberg et Lubart, 1999). L’une des approches psychométriques pour mesurer la créativité implique des tâches de pensée divergente (DT) dans lesquelles les participants sont invités à répondre à un problème donné avec de multiples solutions (Dietrich et Kanso, 2010). Les tests de DT mesurent généralement divers aspects de la créativité, y compris la maîtrise créative, la flexibilité et l’originalité (Torrance, 1974). Ainsi, bien que non synonymes de créativité, les tâches DT fournissent des mesures structurées et objectives de la créativité (Sternberg et Lubart, 1999; Jung et al., 2009). Il a notamment été démontré que les scores des tâches de DT étaient positivement corrélés avec des mesures écologiquement valides de la réussite créative (Carson et al., 2005) ainsi qu’avec une créativité auto-évaluée (Furnham et Bachtiar, 2008).
Des études antérieures ont mis en évidence l’implication du système dopaminergique (DA) dans la créativité (Heilman et al., 2003; Flaherty, 2005; Takeuchi et coll., 2010). Takeuchi et coll. (2010) ont constaté que les différences individuelles de créativité, telles que mesurées par la DT, étaient positivement corrélées avec la matière grise dans les régions du système DA, y compris le cortex préfrontal dorsolatéral, les ganglions basaux bilatéraux, la substantia nigra et la zone ventrale tegmentale. De plus, plusieurs études génétiques ont montré une relation entre la DT et la neurotransmission de la dopamine (Reuter et al., 2006; De Manzano et coll., 2010; Runco et coll., 2011). Par exemple, Reuter et al. (2006) ont constaté que la créativité, mesurée par des tâches DT impliquant à la fois la créativité figurale et verbale, était associée de manière significative aux polymorphismes du gène du récepteur de la dopamine D2 (DRD2). De plus, plusieurs études ont montré que la dopamine corticale était impliquée dans la flexibilité cognitive (Frank, 2005; Cools, 2008; Garcia-Garcia et al., 2010), l’une des principales composantes de DT.
Un gène candidat DA intéressant pour la créativité est le gène du récepteur de la dopamine D4 (DRD4). Le récepteur DRD4 est l’un des cinq récepteurs de la dopamine et joue un rôle important dans la médiation de la signalisation synaptique de la dopamine. Le gène est caractérisé par un nombre variable de 48 paires de bases de répétitions en tandem (VNTR de 48 PB) situées dans la région codante du troisième exon. Le polymorphisme VNTR de 48 pb de DRD4 varie de 2 à 11 répétitions d’un individu à l’autre (Asghari et al., 1995; Cravchik et Goldman, 2000), où la répétition à 4 (4R) est la répétition la plus courante chez les populations caucasiennes et la répétition à 7 (7R) est la deuxième variante la plus courante (Chang et al., 1996; Ding et coll., 2002). Fait intéressant, le 7R a déjà été associé à des comportements créatifs réels, tels que le trait de personnalité à la recherche de nouveauté (Ebstein et al., 1996). En effet, la recherche de nouveauté — la tendance à l’activité exploratoire — est considérée comme l’une des caractéristiques des créateurs (Chavez-Eakle et al., 2006; Drago et coll., 2009). Bien que les preuves d’une association entre DRD4 7R et la recherche de nouveauté soient incohérentes, comme en témoignent plusieurs rapports de méta-analyse (Kluger et al., 2002; Munafò et coll., 2008), de nombreuses études ont trouvé une association significative entre la recherche de nouveauté et l’allèle 7R du DRD4 (Ebstein et al., 1996; Benjamin et coll., 2000; Keltikangas-Järvinen et coll., 2002; Becker et coll., 2005). Roussos et coll. (2009) ont récemment suggéré que les différences dans la mesure de la recherche de nouveauté à l’aide d’échelles d’auto-évaluation pourraient expliquer les résultats incohérents entre le polymorphisme 7R et la recherche de nouveauté. Dans cette optique, il a été constaté à plusieurs reprises que le système DA joue un rôle majeur dans le trait de personnalité de la recherche de nouveauté (Flaherty, 2005; Schweizer, 2006), attestant davantage du rôle potentiel de DRD4 dans la créativité.
Néanmoins, contrairement au rôle du DRD4 48-bp VNTR 7R dans la recherche de nouveauté, des preuves récentes indiquent que le 7R est en fait associé à une altération de la flexibilité cognitive, l’un des aspects majeurs de la créativité (Strobel et al., 2004; Congdon et coll., 2008). La flexibilité implique la capacité de faire une réponse alternative après avoir inhibé avec succès une réponse actuelle. Les résultats obtenus jusqu’à présent sur l’association entre l’inhibition et le DRD4 sont mitigés. Congdon et coll. (2008) ont constaté que les participants avec l’allèle 7R du DRD4 présentent un temps de réaction du signal d’arrêt (SSRT) plus élevé lors d’une tâche go / no-go, reflétant un contrôle inhibiteur plus faible, tandis que Forbes et al. (2009) n’ont pas trouvé un tel effet sur l’Échelle d’impulsivité de Barratt (BRI), un outil d’auto-évaluation pour mesurer l’impulsivité. D’autre part, Colzato et coll. (2010) ont démontré que ces effets mixtes peuvent être dus au fait que des études antérieures abordaient l’impulsivité comme un processus monolithique, alors que l’impulsivité peut en fait être divisée en types fonctionnels et dysfonctionnels selon l’inventaire de l’impulsivité de Dickman (DII; Dickman, 1990). L’impulsivité dysfonctionnelle est la tendance à agir sans prévoyance dans les cas où une telle action est inappropriée, tandis que l’impulsivité fonctionnelle est une tendance similaire mise en œuvre dans des situations appropriées (Colzato et al., 2010). Colzato et coll. (2010) ont constaté que les personnes atteintes de DRD4 7R, qui est associée à des niveaux plus élevés de DA striatal, présentaient une impulsivité dysfonctionnelle plus élevée. Plusieurs études méta-analytiques ont indiqué que l’allèle 7R du DRD4 était un allèle de risque pour le trouble d’hyperactivité avec déficit de l’attention (TDAH; Faraone et al., 2001; DiMaio et coll., 2003), un trouble caractérisé par une impulsivité dysfonctionnelle élevée (Young et al., 2007). De plus, la relation entre le TDAH et la créativité a été étudiée dans plusieurs études (Shaw et Brown, 1990; Healey et Rucklidge, 2006), bien que jusqu’à présent ces études aient donné des résultats peu clairs. Alors que certains ont constaté une corrélation positive entre le TDAH et ses symptômes et la créativité (Healey et Rucklidge, 2006; White et Shah, 2011), d’autres ont signalé une tendance inverse (Funk et al., 1993; Healey et Rucklidge, 2005, 2008). Fait intéressant, le TDAH est associé sur le plan comportemental à des déficiences du contrôle exécutif, y compris le décalage de réglage de l’attention (Boonstra et al., 2005), et la flexibilité (Barkley et coll., 1997; Sergeant et coll., 2002), qui sont toutes deux importantes pour la flexibilité cognitive, une dimension centrale de la créativité (Dietrich, 2004; Durstewitz et Seamans, 2008). Ces résultats indiquent que les personnes avec l’allèle 7R du DRD4 peuvent présenter des niveaux de créativité plus faibles et une flexibilité réduite sur les tâches de DT en particulier. Par conséquent, l’exon III VNTR DRD4 est un candidat biologiquement plausible pour contribuer aux différences individuelles de créativité.
Collectivement, il semble que, d’une part, l’allèle 7R, en tant qu’allèle de risque pour les fonctions exécutives altérées, la flexibilité et le changement de réglage, puisse être associé à une faible créativité. D’autre part, l’association entre l’allèle 7R et une recherche de nouveauté plus élevée indique que les individus avec l’allèle 7R peuvent en fait faire preuve d’une plus grande flexibilité et créativité.
La présente étude a été conçue pour explorer le rôle complexe du DRD4 7R dans la formation partielle de la créativité humaine. Pour mieux caractériser cette relation, nous avons évalué deux types de tests DT (figuraux et verbaux). La notation de chaque tâche a été divisée en fonction des trois dimensions principales de la DT, à savoir la fluidité créative, la flexibilité et l’originalité, et chaque dimension a été analysée séparément pour tester l’hypothèse selon laquelle DRD4 affecte différemment chaque dimension de la créativité.
Méthode
Participants
L’échantillon comprenait 185 étudiants de l’Université de Haïfa en Israël (112 femmes). Tous les participants étaient de race blanche (autodéclarés), avec un âge moyen de 24,5 ans (ET ± 2,1 ans). Les participants ont été recrutés grâce à des annonces affichées à l’université. Tous les participants étaient des bénévoles rémunérés.
Évaluation de la créativité
La créativité a été évaluée à l’aide de deux tests DT, une sous-échelle du sous-test figuratif du test de Torrance de la pensée créative (Torrance, 1974) et la tâche des utilisations alternatives (AUT; Guilford et al., 1978). Les deux tests comprenaient les trois dimensions fondamentales de la DT, à savoir la flexibilité, l’originalité et la fluidité.
Test de Torrance de la Pensée créative (Sous-échelle des cercles)
Les participants ont reçu une page sur laquelle 30 cercles identiques ont été dessinés. On leur a demandé de dessiner autant d’objets significatifs différents que possible dans un délai de 10 min, chaque dessin devant comporter au moins un cercle. La notation comprenait la fluidité (nombre de réponses produites), la flexibilité (nombre de catégories) et l’originalité, calculées en fonction de la notation des réponses originales, comme détaillé dans le guide de notation des tests de Torrance de la pensée créative (Torrance, 1974).
Tâche d’utilisations alternatives (AUT)
Les participants ont reçu une liste de cinq objets communs et ont été invités à énumérer autant d’utilisations alternatives que possible pour chaque objet, dans un délai de 10 min. L’utilisation quotidienne la plus courante était indiquée entre parenthèses. Les objets étaient: chaussure (usage courant: usure à pied); bouton (usage courant: fermeture des objets); crayon (usage courant: dessin ou écriture); pneu (usage courant: roue de voiture); et verre à boire (usage courant: contient du liquide). Seules les réponses qui ne réitéraient pas les utilisations courantes données ont été comptées et incluses. Comme dans le test de Torrance, la notation comprenait la fluidité, la flexibilité et l’originalité. Comme il n’y a pas de lignes directrices pour la notation des réponses originales dans l’AUT, les réponses originales ont été définies comme des réponses statistiquement peu fréquentes selon un pré-test effectué dans notre laboratoire.
Pré-test
Dans le but de créer un critère valide de fréquence de réponse, un groupe de 100 participants en bonne santé qui n’ont pas participé à cette étude a complété l’AUT. Pour chaque objet, une liste de toutes les utilisations possibles a été collectée auprès de tous les participants. Une mesure de fréquence statistique a été calculée à partir de cette liste afin d’évaluer le score d’originalité pour chaque réponse et, par la suite, pour chaque participant. Les réponses ont reçu un score de zéro si 5 % ou plus des participants indiquaient une utilisation donnée, un score de un si entre 2 % et 4,99 % des participants l’indiquaient, et un score de deux si moins de 1,99 % indiquaient l’utilisation. Un score d’originalité moyen a été calculé pour chaque participant en fonction de ces scores statistiques peu fréquents.
Extraction et génotypage de l’ADN
L’ADN a été extrait de 20 ml d’échantillons de rince-bouche à l’aide du kit Master Pure (Epicentre, Madison, WI). Le VNTR DRD4 de 48 pb a été caractérisé par une procédure d’amplification par PCR avec les amorces suivantes: F5′- CTT CCT ACC CTG CCC GCT CAT GCT GCTGCT CTA CTGG-3′ et R5′- ACC ACCACC GGC AGG ACC CTC ATG GCC TTG CGC TC–3. Des réactions de PCR ont été menées en utilisant un mélange maître de 5 µl (Thermo scientific), des amorces de 2 µl (0,5 µM), 0,6 µl Mg / Cl2 (2,5 mM), 0.4 µl de DMSO à 5% et 1 µl d’eau pour un volume total de 9 µl, et 1 µl supplémentaire d’ADN génomique ont été ajoutés au mélange. Toutes les réactions de PCR ont été réalisées sur un Thermocycleur Biometra T1 (Biometra, Güttingem, Allemagne). La condition réactionnelle de PCR était la suivante : étape de préchauffage à 94,0°C pendant 5 min, 34 cycles de dénaturation à 94,0°C pendant 30 s, recuit à 55°C pendant 30 s et extension à 72°C pendant 90 s. La réaction s’est maintenue à 72°C pendant 5 min. Le mélange réactionnel a ensuite été électrophorés sur un gel d’agarose à 3% (AMRESCO) avec du bromure d’éthidium pour cribler les génotypes.
La distribution de la fréquence du génotype était conforme à l’équilibre de Hardy-Weinberg (chi carré = 0,01, valeur de p = 0,92).
L’analyse statistique
Le génotype a été classé en fonction de la présence ou de l’absence de l’allèle 7R du DRD4 (7 contre no7), comme dans les rapports précédents de la littérature (Ding et al., 2002). Les différences de groupe sur les scores de créativité ont été analysées statistiquement à l’aide d’ANOVAs, avec la présence (7) ou l’absence (no7) de l’allèle DRD4 7R comme variable indépendante. Pour examiner les différents aspects de la DT (aisance, flexibilité, originalité), nous avons effectué une analyse multivariée de la variance (MANOVA) séparément pour chaque composant.
Résultats
Pour confirmer que les deux groupes (7, no7) n’étaient pas significativement différents en termes d’âge et de scolarité, nous avons effectué des tests t indépendants pour comparer les deux groupes (voir le tableau 1 pour les moyennes et les écarts types). Cette analyse a révélé que les deux groupes ne différaient pas en termes d’âge ou d’années de scolarité. Des tests non paramétriques (Mann-Whitney) ont révélé des différences significatives dans la distribution de fréquence pour le sexe (Z = 2,11, p < 0,05).
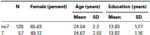
Tableau 1. Données sur l’âge, le sexe et l’éducation pour le génotype DRD4 7R, SD: écart-type.
Les scores moyens des mesures de créativité utilisées dans les deux tâches sont résumés dans le tableau 2.

Tableau 2. Moyenne et Erreur type (SE) des deux tâches de créativité utilisées: AUT et Torrance.
La composante de fluidité
Une ANOVA multivariée indique un effet général du génotype DRD4 7R pour la composante de fluidité, F(2 182) = 3,25, p < 0,05. Compte tenu de l’importance du test global, les effets principaux univariés ont été examinés. Des effets principaux univariés significatifs pour le génotype DRD4 ont été obtenus pour la fluidité AUT F(1 183) = 4,95, p<0.05 (allèle 7R présentant une fluidité inférieure à celle du groupe no7, Figure 1A), mais n’a pas atteint le niveau de signification pour la fluidité de Torrance F (1 183) = 2,95, p = 0,087.
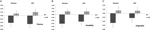
Figure 1. Les scores de créativité sur les tâches AUT et Torrance sont séparés par la fluidité (A), la flexibilité (B) et l’originalité (C). Les moyennes ont été transformées en scores Z.
La composante de flexibilité
L’ANOVA multivariée des mesures de flexibilité a montré un effet général pour le génotype DRD4 7R F (2 182) =4.38, p < 0,05. Des effets principaux univariés significatifs pour le génotype DRD4 ont été obtenus pour la flexibilité AUT F(1 183) = 4,57, p < 0,05 et pour la flexibilité de Torrance F(1 183) = 6,95, p < 0,01. Comme le montre la figure 1B, dans les deux cas, les individus du groupe de l’allèle 7R présentaient une flexibilité inférieure à celle des individus du groupe no7 (tableau 2).
La Composante Originalité
L’ANOVA multivariée des mesures d’originalité n’a présenté aucun effet significatif, F(2 182) = 2,06, p = 0.13, bien que les scores indiquent une originalité inférieure pour l’allèle 7R du DRD4 (voir la figure 1C, tableau 2 pour plus de détails).
Différences entre les sexes
Discussion
Les tâches de DT exigent que les participants apportent plusieurs solutions à un problème donné. Dans les tâches utilisées ici, les participants devaient penser à de nombreuses utilisations possibles alternatives pour les objets du quotidien (dans l’AUT) et dessiner plusieurs dessins incorporant une forme spécifique (cercle) (dans la tâche Torrance). Les résultats présentés ici démontrent que les individus avec l’allèle DRD4 7R présentent une créativité plus faible mesurée par DT. Considérant que l’allèle 7R a été associé à une impulsivité dysfonctionnelle et à une faible inhibition (Congdon et al., 2008; Dreber et coll., 2009), le fait que les individus avec l’allèle 7R présentent une créativité plus faible peut s’expliquer par leur incapacité à supprimer ou à inhiber les réponses évidentes (communes), comme l’indiquent des scores d’originalité plus faibles. Des scores de flexibilité et de fluidité plus faibles peuvent indiquer qu’une fois qu’une réponse commune a été donnée, il devient plus difficile de passer à une nouvelle catégorie (comme l’indiquent les scores de flexibilité faibles), ce qui entraîne moins de réponses (comme l’indique le score de fluidité faible). D’autres preuves de la relation entre la créativité et l’impulsivité peuvent être vues dans l’association de l’allèle 7R avec le TDAH (Li et al., 2006), ainsi qu’avec une impulsivité accrue chez les patients atteints de TDAH (Langley et al., 2004). Fait intéressant, la créativité a été liée au système DA dans le cas de la schizophrénie (Eysenck, 1993). Bien que plusieurs études montrent que les patients atteints de schizophrénie et de personnalité schizotypique présentent une grande créativité (O’Reilly et al., 2001; Folley et Park, 2005), des modèles récents de schizophrénie et de créativité soutiennent une association en forme de U inversé dans laquelle la créativité peut être plus élevée avec un trait schizotypique faible à modéré, mais diminue à mesure que la gravité de la psychopathologie dans la schizophrénie augmente (Tsakanikos et Claridge, 2005; Stoneham et Coughtrey, 2009; Nelson et Rawlings, 2010). En effet, dans une étude récente Jaracz et al. (2012) ont rapporté que les patients atteints de schizophrénie présentaient une créativité diminuée par rapport aux témoins. En outre, les auteurs ont rapporté que la créativité altérée était associée à une faible flexibilité cognitive et à une impulsivité faibles, reflétées par leurs faibles scores au test de tri des cartes du Wisconsin (WCST). Ces résultats sont conformes aux recherches antérieures montrant que les patients atteints de schizophrénie chronique présentent une créativité plus faible, ce qui, dans le cas de la maîtrise créative, était médié par des scores sur les tâches de contrôle exécutif et, dans le cas de l’originalité, ne l’était pas (Abraham et al., 2007). Collectivement, alors que l’impulsivité fonctionnelle légère peut améliorer la créativité chez les patients présentant des traits schizotypiques, en particulier en générant des idées qui diffèrent des exemples donnés (Abraham et Windmann, 2008), l’impulsivité dysfonctionnelle observée dans la schizophrénie sévère peut atténuer les capacités créatives.
Les résultats de la présente étude confirment la relation entre la créativité et les variantes des polymorphismes génétiques DA (Reuter et al., 2006; Runco et coll., 2011). De plus, les résultats trouvés dans notre échantillon indiquent que le principal effet de l’allèle DRD4-7R sur la créativité peut être dû à son impact sur la flexibilité. Plusieurs études ont établi un lien entre la flexibilité cognitive et les voies DA (Ashby et Isen, 1999; Dreisbach et Goschke, 2004; Müller et al., 2007). Il a été postulé que l’activation des récepteurs de type D2 (c.-à-d., D2, D3 et D4) diminue l’inhibition GABAergique et facilite l’activation de représentations multiples, augmentant ainsi la flexibilité comme le montrent les capacités de décalage de réglage (Müller et al., 2007; Seamans et Robbins, 2010). De plus, les récepteurs de type D2 ont été liés à une intégration flexible de nouvelles informations (Durstewitz et Seamans, 2008). Dans cette optique, Reuter et al. (2006) ont trouvé une association entre le récepteur de la dopamine D2 et la créativité mesurée par un indice composite, bien qu’il ne soit pas possible de déterminer si la flexibilité a joué un rôle majeur dans ces résultats. De plus, Runco et coll. (2011) ont signalé des différences dans la fluidité créative entre les porteurs de plusieurs gènes dopaminergiques, parmi lesquels le DRD4. Cependant, il n’est pas clair d’après leur rapport quel groupe présente de plus grandes capacités créatives. Il est important de noter que, contrairement aux résultats présentés ici, Runco et al. (2011) n’ont pas trouvé d’association entre les mesures de flexibilité mesurées par les tâches DT et DRD4. Cette incohérence peut s’expliquer par des différences dans les populations utilisées et la taille des groupes ou par des différences dans les fréquences des allèles au sein de la population échantillonnée. De plus, il existe des preuves suggérant que l’association entre la dopamine et la DT peut ne pas être linéaire (Akbari Chermahini et Hommel, 2010, 2012), indiquant qu’une interaction entre plusieurs gènes DA peut influencer les résultats individuels de chaque variabilité génétique dans le contexte de la DT. En outre, plusieurs études ont mis en évidence le fait que les voies de DA peuvent être sensibles au genre (Tammimäki et Männistö, 2011; Wang et al., 2012). Zhong et coll. (2010), par exemple, ont constaté que le genre modulait l’association entre le polymorphisme de l’exonon3 DRD4 et le sens de l’équité. Dans la présente étude, nous n’avons pas trouvé d’effet d’interaction entre le DRD4-7R et le sexe, ce qui indique que le sexe n’a pas d’effet modulateur sur l’association entre le DRD4 et la créativité dans notre échantillon. En outre, les résultats présentés ici indiquent une absence de différences entre les sexes dans la créativité, et en particulier dans la DT (pour examen, voir Baer et Kaufman, 2008).
En conclusion, dans la présente étude, nous suggérons que l’association trouvée entre DRD4 et la créativité est principalement influencée par la flexibilité. Les résultats de la présente étude sont conformes aux résultats précédents indiquant l’implication du système DA dans la créativité et s’ajoutent à l’accumulation de connaissances suggérant des influences génétiques dans la créativité.
Il y a plusieurs limites de l’étude qui doivent être reconnues. Premièrement, l’étude actuelle n’a examiné que l’association entre une variabilité génétique dans le système DA — le DRD4 — et le DT. Cependant, il existe d’autres polymorphismes importants dans le système DA striatal qui pourraient être associés à la DT et peuvent contribuer à la fois de manière autonome et dans les interactions gène-gène. L’un de ces polymorphismes est le transporteur de dopamine (DAT1), qui s’est avéré associé à l’inhibition du retour (Colzato et al., 2010) ainsi qu’avec des mesures de l’impulsivité (Gizer et Waldman, 2012). De plus, comme mentionné ci-dessus, une autre possibilité qui n’a pas été explorée ici est les effets possibles de l’interaction gène-gène sur la DT. Akbari Chermahini et Hommel (2010) ont rapporté une association non linéaire entre un marqueur de dopamine striatale et la DT, indiquant que la relation peut être plus complexe, de sorte qu’un polymorphisme d’un gène peut avoir un impact sur la DT par la médiation d’un autre polymorphisme génétique. Ainsi, les études futures devraient envisager d’examiner l’interaction entre ces deux polymorphismes dans le contexte de la flexibilité et de la DT.
Déclaration sur les conflits d’intérêts
Les auteurs déclarent que la recherche a été menée en l’absence de relations commerciales ou financières pouvant être interprétées comme un conflit d’intérêts potentiel.
Akbari Chermahini, S., et Hommel, B. (2010). Le (b) lien entre la créativité et la dopamine: les taux de clignotement spontanés des yeux prédisent et dissocient la pensée divergente et convergente. Cognition 115, 458-465. doi: 10.1016 /j. cognition.2010.03.007
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Cools, R. (2008). Rôle de la dopamine dans le contrôle motivationnel et cognitif du comportement. Neuroscientifique 14, 381-395. doi: 10.1177 / 1073858408317009
Résumé Pubmed / Texte Intégral Pubmed / Texte intégral CrossRef
Frank, M. J. (2005). Modulation dynamique de la dopamine dans les ganglions de la base: un compte neurocomputationnel des déficits cognitifs dans le parkinsonisme médicamenteux et non médicamenteux. J. Cogn. Neurosci. 17, 51–72. doi: 10.1162/0898929052880093
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Healey, D., et Rucklidge, J. J. (2006). Une enquête sur la relation entre la symptomatologie du TDAH, la créativité et le fonctionnement neuropsychologique chez les enfants. Neuropsychol infantile. 12, 421–438. doi: 10.1080/09297040600806086
Résumé Pubmed/Texte Intégral Pubmed/Texte intégral CrossRef
Runco, M.A., et Richards, R. (1997). Créativité Éminente, Créativité Quotidienne et Santé. Greenwich, CT : Ablex.
Torrance, E. (1974). Les différences ne sont pas des déficits. Enseigner. Coll. Rec. 75, 471–487.