- Bevezetés
- módszer
- résztvevők
- A kreativitás értékelése
- A kreatív gondolkodás Torrance tesztje (körök Alskála)
- alternatív felhasználási feladat (aut)
- Elővizsgálat
- DNS-extrakció és genotipizálás
- statisztikai elemzés
- eredmények
- a folyékony komponens
- a rugalmassági intézkedések
- az eredetiség komponens
- nemek közötti különbségek
- Vita
- összeférhetetlenségi nyilatkozat
Bevezetés
a kreatív megismerés fontos szerepet játszik a művészetekben, a találmányokban és az innovációban, valamint a mindennapi életben (Runco and Richards, 1997). Bár a kreativitást egyedülálló emberi képességnek tekintik, a spontán kreatív viselkedés (pl., új eszközök létrehozása a főemlősök és a madarak között) kimutatták, hogy nem embereknél is előfordulnak (Byrne és Bates, 2007), tovább igazolva a kreativitás mély evolúciós és biológiai gyökereit. A kreativitás az újszerű (eredeti, ritka és váratlan) és alkalmas (adaptív és hasznos a feladat korlátai szerint) válaszok előállításának képessége (Sternberg and Lubart, 1999). A kreativitás mérésének egyik pszichometriai megközelítése a divergens gondolkodás (DT) feladatait foglalja magában, amelyekben a résztvevőket arra kérik, hogy válaszoljanak egy adott problémára több megoldással (Dietrich and Kanso, 2010). A DT tesztjei általában a kreativitás különböző aspektusait mérik, beleértve a kreatív folyékonyságot, a rugalmasságot és az eredetiséget (Torrance, 1974). Így, bár nem szinonimája a kreativitásnak, a DT feladatok strukturált és objektív méréseket nyújtanak a kreativitásról (Sternberg and Lubart, 1999; Jung et al., 2009). Nevezetesen kimutatták, hogy a DT feladatok pontszáma pozitívan korrelál a kreatív teljesítmény ökológiailag érvényes intézkedéseivel (Carson et al., 2005), valamint önértékelt kreativitással (Furnham and Bachtiar, 2008).
korábbi tanulmányok rámutattak a dopaminerg (DA) rendszer részvételére a kreativitásban (Heilman et al., 2003; Flaherty, 2005; Takeuchi et al., 2010). Takeuchi et al. (2010) megállapította, hogy a kreativitás egyéni különbségei a DT által mérve pozitívan korrelálnak a szürkeállománysal A DA rendszer régióiban, beleértve a dorsolaterális prefrontális kérget, a bilaterális bazális ganglionokat, a substantia nigra-t és a tegmentális ventrális területet. Ezenkívül számos genetikai vizsgálat kimutatta a DT és a dopamin neurotranszmisszió közötti kapcsolatot (Reuter et al., 2006; De Manzano et al., 2010; Runco et al., 2011). Például Reuter et al. (2006) megállapította, hogy a kreativitás, mind a figurális, mind a verbális kreativitást magában foglaló DT feladatokkal mérve, jelentősen összefügg a dopamin D2 receptor gén (DRD2) polimorfizmusaival. Sőt, számos tanulmány megállapította, hogy a kortikális dopamin részt vesz a kognitív rugalmasságban (Frank, 2005; Cools, 2008; Garcia-Garcia et al., 2010), a DT egyik fő összetevője.
a kreativitás egyik érdekes DA jelölt génje a dopamin D4 receptor gén (DRD4). A DRD4 receptor az öt dopamin receptor egyike, és fontos szerepet játszik a szinaptikus dopamin jelátvitel közvetítésében. A gént 48 bázispár változó számú tandem ismétlés jellemzi (48 bp VNTR), amely a harmadik exon kódoló régiójában található. A DRD4 48-bp VNTR polimorfizmus 2-11 ismétlődés között változik az egyének között (Asghari et al. 1995; Cravchik and Goldman, 2000), ahol a 4-ismétlés (4R) a leggyakoribb ismétlés a kaukázusi populációkban, a 7-ismétlés (7R) pedig a második leggyakoribb változat (Chang et al., 1996; Ding et al., 2002). Érdekes módon a 7R-t korábban összefüggésbe hozták a valós kreatív viselkedéssel, például az újdonságot kereső személyiségvonással (Ebstein et al., 1996). Valóban, újdonság-kereső-a tendencia a feltáró tevékenység-úgy gondolják, hogy az egyik jellemzője a kreatív emberek (Chavez-Eakle et al., 2006; Drago et al., 2009). Bár a DRD4 7R és az újdonságkeresés közötti összefüggés bizonyítékai következetlenek, amint azt számos metaanalízis-jelentés is bizonyítja (Kluger et al., 2002; Munaf stb., 2008), számos tanulmány jelentős összefüggést talált az újdonságkeresés és a DRD4 7R allélja között (Ebstein et al., 1996; Benjamin et al., 2000; Keltikangas-J Enterprvinen et al., 2002; Becker et al., 2005). Roussos et al. (2009) nemrégiben azt javasolta, hogy az újdonságkeresés önjelentési skálákkal történő mérésének különbségei magyarázhatják a 7R polimorfizmus és az újdonságkeresés közötti következetlen eredményeket. Ezzel összhangban ismételten megállapítást nyert, hogy a DA rendszer jelentős szerepet játszik az újdonságkeresés személyiségjegyében (Flaherty, 2005; Schweizer, 2006), tovább tanúsítva a DRD4 potenciális szerepét a kreativitásban.
ennek ellenére, ellentétben a DRD4 48-bp VNTR 7R szerepével az újdonságkeresésben, a legújabb bizonyítékok azt mutatják, hogy a 7R valójában a kognitív rugalmasság károsodásával jár, amely a kreativitás egyik fő szempontja (Strobel et al., 2004; Congdon et al., 2008). A rugalmasság magában foglalja azt a képességet, hogy alternatív választ adjon az aktuális válasz sikeres gátlása után. A gátlás és a DRD4 közötti összefüggésre vonatkozó eddigi eredmények vegyesek. Congdon et al. (2008) megállapította, hogy a DRD4 7R alléljával rendelkező résztvevők magasabb stop-jel reakcióidőt (SSRT) mutatnak egy go/no-go feladaton, ami a gyengébb gátló kontrollt tükrözi, míg Forbes et al. (2009) nem talált ilyen hatást a Barratt impulzivitás skála (BIS), egy önjelentési eszköz az impulzivitás mérésére. Másrészről, Colzato et al. (2010) kimutatta, hogy ezek a vegyes hatások annak a ténynek tudhatók be, hogy a korábbi tanulmányok az impulzivitást monolitikus folyamatként kezelték, míg az impulzivitás valójában funkcionális és diszfunkcionális típusokra osztható a Dickman Impulzivitási leltár (DII; Dickman, 1990) szerint. A diszfunkcionális impulzivitás az a tendencia, hogy előrelátás nélkül cselekszünk azokban az esetekben, amikor az ilyen cselekvés nem megfelelő, míg a funkcionális impulzivitás hasonló tendencia, amelyet megfelelő helyzetekben hajtanak végre (Colzato et al., 2010). Colzato et al. (2010) megállapította, hogy a DRD4 7R-vel rendelkező egyének, amelyek a striatális DA magasabb szintjéhez kapcsolódnak, magasabb diszfunkcionális impulzivitást mutattak. Számos meta-analitikai tanulmány rámutatott a DRD4 7R alléljára, mint a figyelemhiányos hiperaktivitási rendellenesség kockázati alléljára (ADHD; Faraone et al., 2001; DiMaio et al., 2003), egy olyan rendellenesség, amelyet magas diszfunkcionális impulzivitás jellemez (Young et al., 2007). Ezenkívül az ADHD és a kreativitás közötti kapcsolatot számos tanulmányban vizsgálták (Shaw és Brown, 1990; Healey és Rucklidge, 2006), bár eddig ezek a tanulmányok nem egyértelmű eredményeket hoztak. Míg egyesek úgy találták, hogy az ADHD és tünetei pozitívan korrelálnak a kreativitással (Healey and Rucklidge, 2006; White and Shah, 2011), mások ellentétes tendenciáról számoltak be (Funk et al., 1993; Healey and Rucklidge, 2005, 2008). Érdekes módon az ADHD viselkedésileg összefügg a végrehajtó kontroll károsodásával, beleértve a figyelmi set-shiftinget (Boonstra et al., 2005) és a rugalmasság (Barkley et al., 1997; őrmester et al., 2002), mindkettő fontos a kognitív rugalmasság, a kreativitás központi dimenziója szempontjából (Dietrich, 2004; Durstewitz and Seamans, 2008). Ezek az eredmények azt mutatják, hogy a DRD4 7R alléljával rendelkező egyének alacsonyabb szintű kreativitást és csökkent rugalmasságot mutathatnak a DT feladatokban. Ezért a DRD4 exon III VNTR biológiailag elfogadható jelölt a kreativitás egyéni különbségeihez való hozzájáruláshoz.
összességében úgy tűnik, hogy egyrészt a 7R allél, mint a csökkent végrehajtó funkciók, a rugalmasság és a halmazeltolódás kockázati allélja, alacsony kreativitással járhat. Másrészt a 7R allél és a magasabb újdonságkeresés közötti kapcsolat azt jelzi, hogy a 7R alléllel rendelkező egyének nagyobb rugalmasságot és kreativitást mutathatnak.
a jelenlegi tanulmány célja a DRD4 7R összetett szerepének feltárása az emberi kreativitás részleges alakításában. Ennek a kapcsolatnak a jobb jellemzése érdekében kétféle DT tesztet értékeltünk (figurális és verbális). Az egyes feladatok pontozását a DT három fő dimenziója szerint osztották fel, nevezetesen a kreatív folyékonyság, a rugalmasság és az eredetiség, és minden dimenziót külön-külön elemeztek annak a hipotézisnek a tesztelésére, hogy a DRD4 a kreativitás minden dimenzióját eltérő módon befolyásolja.
módszer
résztvevők
a minta 185 hallgatót tartalmazott az Izraeli Haifa Egyetemen (112 nő). Minden résztvevő kaukázusi volt (önjelentés szerint), átlagéletkora 24,5 volt (SD 2,1). A résztvevőket az egyetemen közzétett bejelentések révén toborozták. Minden résztvevő fizetett önkéntes volt.
A kreativitás értékelése
a kreativitást két DT teszt segítségével értékelték, a Torrance kreatív gondolkodás tesztjének figurális alpróbájából (Torrance, 1974) és az alternatív felhasználási feladatból (aut; Guilford et al., 1978). Mindkét teszt magában foglalta a DT három alapvető dimenzióját, nevezetesen a rugalmasságot, az eredetiséget és a folyékonyságot.
A kreatív gondolkodás Torrance tesztje (körök Alskála)
a résztvevők kaptak egy oldalt, amelyen 30 azonos kört rajzoltak. Arra kérték őket, hogy rajzoljanak annyi különböző értelmes tárgyat, amennyire csak lehetséges, 10 perces határidőn belül, ahol minden rajznak tartalmaznia kell legalább egy kört. A pontozás magában foglalta a folyékonyságot (az előállított válaszok számát), a rugalmasságot (a kategóriák számát) és az eredetiséget, az eredeti válaszok pontozása alapján számítva, amint azt a Torrance Tests of Creative Thinking pontozási útmutató (Torrance, 1974) részletezi.
alternatív felhasználási feladat (aut)
a résztvevők öt közös objektum listáját kapták, és felkérték, hogy sorolják fel az egyes objektumokhoz a lehető legtöbb alternatív felhasználást, 10 perces határidőn belül. A leggyakoribb mindennapi használatot zárójelben jeleztük. A tárgyak a következők voltak: cipő (gyakori használat: kopás gyalog); gomb (gyakori használat: záró dolgok); ceruza (gyakori használat: rajz vagy írás); gumiabroncs (gyakori használat: autó kerék); és ivópohár (gyakori használat: folyadékot tartalmaz). Csak azokat a válaszokat számoltuk És vettük figyelembe, amelyek nem ismételték meg az adott közös felhasználást. A Torrance teszthez hasonlóan a pontozás magában foglalta a folyékonyságot, a rugalmasságot és az eredetiséget. Mivel az Aut-ban nincsenek irányelvek az eredeti válaszok pontozására, az eredeti válaszokat statisztikailag ritka válaszként határoztuk meg a laboratóriumunkban végzett előzetes vizsgálat szerint.
Elővizsgálat
a válaszfrekvencia érvényes kritériumának létrehozása céljából egy 100 egészséges résztvevőből álló csoport, akik nem vettek részt ebben a vizsgálatban, befejezték az AUT-t. Minden objektumhoz összegyűjtötték az összes lehetséges felhasználás listáját az összes résztvevőtől. E lista alapján statisztikai ritkaság-mérést számítottak ki az egyes válaszok eredetiség-pontszámának értékelése érdekében, majd ezt követően minden résztvevő számára. A válaszok nulla pontszámot kaptak, ha a résztvevők legalább 5% – a felsorolt egy adott felhasználást, egy pontszámot, ha a résztvevők 2% és 4,99% között szerepel, és két pontszámot, ha kevesebb, mint 1,99% sorolta fel a felhasználást. Átlagos eredetiség pontszámot számítottak ki minden résztvevő számára e statisztikai ritkasági pontszámok alapján.
DNS-extrakció és genotipizálás
a DNS-t 20 ml szájvízmintából vontuk ki a Master Pure kit (Epicentre, Madison, WI) segítségével. A DRD4 48-bp VNTR – t PCR amplifikációs eljárással jellemeztük a következő primerekkel: F5′ – CTT CCT ACC CTG CCC GCT CAT GCT GCTGCT CTA CTGG – 3′ és R5′ – ACC ACCACC GGC AGG ACC CTC ATG GCC TTG CGC TC-3. A PCR-reakciókat 5 fő (Thermo scientific), 2 fő (0,5 fő), 0,6 fő (mg/Cl2) (2,5 mM), 0 fő alkalmazásával végeztük.4 db 5% DMSO-t és 1 db vízmennyiséget összesen 9 db teljes térfogatra, és további 1 db genomi DNS-t adtunk az elegyhez. Az összes PCR-reakciót Biometra T1 Termocikloron végeztük (Biometra, G ons, Németország). A PCR-reakció állapota a következő volt: előmelegítési lépés 94,0 6 perc alatt 5 perc alatt, 34 ciklus denaturálás 94,0 x 30 másodperc alatt, újrazárás 55 x 30 másodperc alatt 52 x XNUMX perc alatt, majd meghosszabbítás 72 x 90 másodperc alatt. a reakció 72 x 5 percig tartott. A reakcióelegyet ezután 3% – os agarózgélen (AMRESCO) etidium-bromiddal elektroforézist végeztünk a genotípusok szűrésére.
a genotípus gyakoriságának eloszlása a Hardy-Weinberg egyensúly (chi-négyzet = 0,01, p érték = 0,92).
statisztikai elemzés
a genotípust a DRD 7R alléljának jelenléte vagy hiánya szerint osztályozták4 (7 vs. no7), a szakirodalomban szereplő korábbi jelentésekkel összhangban (Ding et al., 2002). A kreativitási pontszámok csoportbeli különbségeit statisztikailag anovák alkalmazásával elemeztük, független változóként a DRD4 7R allél jelenléte (7) vagy hiánya (no7). A DT különböző aspektusainak (folyékonyság, rugalmasság, eredetiség) vizsgálatához többváltozós varianciaanalízist (MANOVA) végeztünk külön-külön az egyes komponensekre.
eredmények
annak igazolására, hogy a két csoport (7, no7) nem különbözött szignifikánsan az életkor és az oktatás szempontjából, független t-teszteket végeztünk a két csoport összehasonlítására (az átlagokat és a szórásokat lásd az 1.táblázatban). Ez az elemzés feltárta, hogy a két csoport nem különbözött életkor vagy iskolai végzettség szempontjából. A nem parametrikus tesztek (Mann–Whitney) szignifikáns különbségeket tártak fel a nemek gyakoriságának eloszlásában (Z = 2,11, p < 0,05).
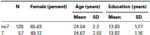
1. táblázat. A DRD4 7R genotípus életkorra, nemre és iskolai végzettségre vonatkozó adatai, SD: szórás.
a két feladat során alkalmazott kreativitási intézkedések átlagos pontszámait a 2.táblázat foglalja össze.

2. táblázat. A két alkalmazott kreativitási feladat eszköze és Standard hibája (SE) : AUT és Torrance.
a folyékony komponens
egy többváltozós ANOVA a DRD4 7R genotípus általános hatását jelezte a folyékony komponensre, F(2,182) = 3,25, p < 0,05. Tekintettel a teljes teszt jelentőségére, egyváltozós fő hatásokat vizsgáltunk. A DRD4 genotípusra vonatkozó jelentős egyváltozós fő hatásokat aut folyékonyság F (1,183) = 4,95, p < 0.05 (7R allél, amely alacsonyabb folyékonyságot mutat a no7 csoporthoz képest, 1a ábra), de nem érte el a Torrance folyékonyság szignifikanciaszintjét F(1,183) = 2,95, p = 0,087.
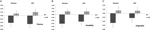
1. ábra. A kreativitás pontszáma az AUT és Torrance feladatokon, a folyékonyságra (A), a rugalmasságra (B) és az eredetiségre (C) elkülönítve. Az eszközöket Z pontszámokká alakították át.
a rugalmassági intézkedések
többváltozós Anova rugalmassági összetevője általános hatást mutatott a DRD4 7R F genotípusra(2182) = 4.38, p < 0,05. A DRD4 genotípusra vonatkozó jelentős egyváltozós fő hatásokat az AUT flexibility F(1183) = 4,57, p < 0,05 és a Torrance flexibility F(1183) = 6,95, p < 0,01 esetében érték el. Amint az 1b. ábrán látható, mindkét esetben a 7R allélcsoport egyedei alacsonyabb rugalmasságot mutattak, mint az no7 csoport egyedei (2.táblázat).
az eredetiség komponens
az eredetiségmérések többváltozós ANOVA-ja nem mutatott szignifikáns hatást, F(2182) = 2,06, p = 0.13, bár a pontszámok a DRD4 7R alléljának alacsonyabb eredetiségét mutatták (a részletekért lásd az 1C.ábrát, a 2. táblázatot).
nemek közötti különbségek
Vita
a DT feladatok megkövetelik a résztvevőktől, hogy több megoldást nyújtsanak egy adott problémára. Az itt alkalmazott feladatokban a résztvevőknek számos alternatív lehetséges felhasználási módot kellett gondolniuk a mindennapi tárgyakra (az AUT-ban), és több rajzot kellett rajzolniuk, amelyek egy adott alakot (kört) tartalmaznak (a Torrance-feladatban). Az itt bemutatott eredmények azt mutatják, hogy a DRD4 7R alléllel rendelkező egyének alacsonyabb kreativitást mutatnak a DT-vel mérve. Figyelembe véve, hogy a 7R allél diszfunkcionális impulzivitással és gyenge gátlással jár (Congdon et al., 2008; Dreber et al., 2009), az a tény, hogy a 7R alléllel rendelkező egyének alacsonyabb kreativitást mutatnak, azzal magyarázható, hogy képtelenek elnyomni vagy gátolni a nyilvánvaló (gyakori) válaszokat, amint azt az alacsonyabb eredetiségi pontszámok jelzik. Az alacsonyabb rugalmassági és folyékonysági pontszámok azt jelezhetik, hogy ha egyszer közös választ adtak, nehezebbé válik az új kategóriára való áttérés (amint azt az alacsony rugalmassági pontszámok jelzik), ami kevesebb válaszhoz vezet (amint azt az alacsony folyékonysági pontszám jelzi). A kreativitás és az impulzivitás közötti kapcsolat további bizonyítékai láthatók a 7R allél ADHD-vel való összefüggésében (Li et al., 2006), valamint az ADHD-s betegek fokozott impulzivitásával (Langley et al., 2004). Érdekes módon a kreativitás összekapcsolódott a DA rendszerrel skizofrénia esetén (Eysenck, 1993). Bár számos tanulmány azt mutatja, hogy a skizofréniában és a skizotípusos személyiségben szenvedő betegek nagy kreativitást mutatnak (O ‘ Reilly et al. 2001; Folley and Park, 2005), a skizofrénia és a kreativitás legújabb modelljei egy fordított U-alakú asszociációt támogatnak, amelyben a kreativitás magasabb lehet alacsony-közepes skizotípusos vonással, de csökken, mivel a skizofrénia pszichopatológiájának súlyossága növekszik (Tsakanikos and Claridge, 2005; Stoneham and Coughtrey, 2009; Nelson and Rawlings, 2010). Valóban, egy nemrégiben készült tanulmányban Jaracz et al. (2012) arról számolt be, hogy a skizofréniában szenvedő betegek csökkent kreativitást mutatnak a kontrollokhoz képest. Továbbá a szerzők arról számoltak be, hogy a csökkent kreativitás alacsony kognitív rugalmassággal és impulzivitással társult, amit a Wisconsin Card Válogatás teszt (WCST) alacsony pontszáma tükröz. Ezek az eredmények összhangban vannak a korábbi kutatásokkal, amelyek azt mutatják, hogy a krónikus skizofréniában szenvedő betegek alacsonyabb kreativitást mutatnak, amelyet a kreatív folyékonyság esetén a végrehajtó ellenőrzési feladatok pontszáma közvetített, az eredetiség esetében pedig nem (Abraham et al., 2007). Összességében, míg az enyhe funkcionális impulzivitás javíthatja a kreativitást a skizotípusos tulajdonságokkal rendelkező betegek körében, különösen a bemutatott példáktól eltérő ötletek generálásában (Abraham and Windmann, 2008), a súlyos skizofréniában megfigyelt diszfunkcionális impulzivitás tompíthatja a kreatív képességeket.
a jelenlegi tanulmány eredményei alátámasztják a kreativitás és a DA genetikai polimorfizmusok variánsai közötti kapcsolatot (Reuter et al., 2006; Runco et al., 2011). Sőt, a mintánkban talált eredmények azt mutatják, hogy a DRD4-7R allél kreativitásra gyakorolt fő hatása a rugalmasságra gyakorolt hatásának tudható be. Számos tanulmány kapcsolta össze a kognitív rugalmasságot a DA pathways-szel (Ashby and Isen, 1999; Dreisbach and Goschke, 2004; M)., 2007). Feltételezték, hogy a D2-szerű receptorok (azaz a D2, D3 és D4) aktiválása csökkenti a GABAerg gátlást és megkönnyíti a többszörös reprezentációk aktiválását, ezáltal növelve a rugalmasságot, amelyet a set-shifting képességek mutatnak (M)., 2007; Seamans and Robbins, 2010). Ezenkívül a D2-szerű receptorok összekapcsolódtak az új információk rugalmas integrációjával (Durstewitz and Seamans, 2008). Ezzel összhangban, Reuter et al. (2006) összefüggést talált a D2 dopaminreceptor és a kreativitás között, amelyet összetett index alapján mértek, bár nem lehet meghatározni, hogy a rugalmasság fontos szerepet játszott-e ezekben az eredményekben. Továbbá, Runco et al. (2011) a kreatív folyékonyság különbségeiről számolt be több dopamin gén hordozói között, köztük a DRD4. Jelentésükből azonban nem derül ki, melyik csoport mutat nagyobb kreatív képességeket. Fontos megjegyezni, hogy az itt bemutatott eredményekkel ellentétben Runco et al. (2011) nem talált összefüggést a DT feladatokkal mért rugalmassági intézkedések és a DRD4 között. Ez az inkonzisztencia magyarázható az alkalmazott populációk és a csoportméretek különbségeivel, vagy a mintavételezett populáción belüli allélfrekvenciák különbségeivel. Továbbá bizonyíték van arra, hogy a dopamin és a DT közötti kapcsolat nem lehet lineáris (Akbari Chermahini and Hommel, 2010, 2012), ami azt jelzi, hogy a több DA gén közötti kölcsönhatás befolyásolhatja az egyes genetikai variabilitás egyedi eredményeit a DT összefüggésében. Ezenkívül számos tanulmány rámutatott arra a tényre, hogy a DA pathways nemi szempontból érzékeny lehet (Tammim Multinnist, 2011; Wang et al., 2012). Zhong et al. (2010) például megállapította, hogy a nemek modulálták a DRD4 exon3 polimorfizmus és a méltányosság érzését. A jelenlegi tanulmányban nem találtunk interakciós hatást a DRD4-7R és a nemek között, ami azt jelzi, hogy a nemnek nincs moduláló hatása a DRD4 és a kreativitás közötti kapcsolatra a mintánkban. Továbbá az itt bemutatott eredmények azt mutatják, hogy a kreativitás és különösen a DT terén nincsenek nemek közötti különbségek (lásd: Baer and Kaufman, 2008).összefoglalva, a jelenlegi tanulmányban azt javasoljuk, hogy a DRD4 és a kreativitás közötti összefüggést elsősorban a rugalmasság befolyásolja. A jelenlegi tanulmány eredményei összhangban vannak a DA rendszer kreativitásban való részvételére utaló korábbi eredményekkel, és hozzáadják a felhalmozódó tudást, ami a kreativitás genetikai befolyásolására utal.
a tanulmánynak számos korlátja van, amelyeket el kell ismerni. Először is, a jelenlegi tanulmány csak a Da rendszer egyik genetikai variabilitása—a DRD4—és a DT közötti összefüggést vizsgálta. Vannak azonban más fontos polimorfizmusok a striatális DA rendszerben, amelyek összefüggésbe hozhatók a DT-vel, és hozzájárulhatnak mind önálló módon, mind a gén-gén kölcsönhatásokban. Az egyik ilyen polimorfizmus a dopamin transzporter (DAT1), amelyről kimutatták, hogy összefügg a visszatérés gátlásával (Colzato et al., 2010), valamint az impulzivitás mértékével (Gizer and Waldman, 2012). Továbbá, amint fentebb említettük, egy másik lehetőség, amelyet itt nem vizsgáltunk, a lehetséges gén-gén interakciós hatások a DT-re. Akbari Chermahini and Hommel (2010) nemlineáris összefüggésről számolt be a striatális dopamin és a DT markere között, jelezve, hogy a kapcsolat összetettebb lehet, így egy gén egyik polimorfizmusa befolyásolhatja a DT-t egy másik gén polimorfizmusának közvetítésével. Ezért a jövőbeni tanulmányoknak fontolóra kell venniük e két polimorfizmus kölcsönhatásának vizsgálatát a rugalmasság és a DT összefüggésében.
összeférhetetlenségi nyilatkozat
a szerzők kijelentik, hogy a kutatást olyan kereskedelmi vagy pénzügyi kapcsolatok hiányában végezték, amelyek potenciális összeférhetetlenségnek tekinthetők.
Akbari Chermahini, S., and Hommel, B. (2010). A (b) kapcsolat a kreativitás és a dopamin között: a spontán szemvillanási Arány megjósolja és disszociálja a divergens és konvergens gondolkodást. Megismerés 115, 458-465. doi: 10.1016 / j. megismerés.2010.03.007
Pubmed absztrakt / Pubmed teljes szöveg / CrossRef teljes szöveg
Cools, R. (2008). A dopamin szerepe a viselkedés motivációs és kognitív kontrolljában. Idegtudós 14, 381-395. doi: 10.1177/1073858408317009
Pubmed absztrakt / Pubmed teljes szöveg / CrossRef teljes szöveg
Frank, M. J. (2005). Dinamikus dopamin moduláció a bazális ganglionokban: a gyógyszeres és nem gyógyszeres parkinsonizmus kognitív deficitjeinek neurokomputációs beszámolója. J. Cogn. Neurosci. 17, 51–72. doi: 10.1162 / 0898929052880093
Pubmed absztrakt / Pubmed teljes szöveg / CrossRef teljes szöveg
Healey, D., and Rucklidge, J. J. (2006). Az ADHD tünetei, a kreativitás és a neuropszichológiai működés közötti kapcsolat vizsgálata gyermekeknél. Gyermek Neuropszichol. 12, 421–438. doi: 10.1080 / 09297040600806086
Pubmed absztrakt / Pubmed teljes szöveg / CrossRef teljes szöveg
Runco, Ma, and Richards, R. (1997). Kiemelkedő kreativitás, mindennapi kreativitás és egészség. Greenwich, CT: Ablex.
Torrance, E. (1974). A különbségek nem hiányok. Taníts. Coll. Rec. 75, 471–487.