- Bevezetés
- anyagok és módszerek
- növényi anyagok és Növekedési állapot
- PSI és PSII mérések
- Elektrokróm Shift analízis
- Fotoinhibitor kezelések
- statisztikai elemzés
- eredmények
- Vita
- a CEF stimuláció szerepe gyenge fényviszonyok mellett
- A P700 oxidációs arányának változása gyenge fényviszonyok mellett nem kapcsolódik a pmf-hez
- az ETRII szabályozza a P700 oxidációs arányt gyenge fényviszonyok mellett
- következtetés
- szerzői hozzájárulások
- finanszírozás
- összeférhetetlenségi nyilatkozat
Bevezetés
bár a fény a hajtóereje a fotoszintézis, nagy fény okozhat jelentős fotoinhibition photosystem II (PSII) a levelek árnyék-létrehozó növények (Kitao et al., 2000; Barth et al., 2001; Krause et al., 2004; Huang et al., 2015b, 2016b, c). A fotoinhibitált PSII azonban gyenge fényviszonyok mellett gyorsan helyreáll (He and Chow, 2003; Zhang és Scheller, 2004, Allakhverdiev et al., 2005), a D1 fehérje gyors forgalmának köszönhetően (Aro et al., 1993; Zhang and Scheller, 2004; Allakhverdiev et al., 2005). A PSII javulásának sebessége csökkent az ATP szintézisének gátlásakor, akár PSI, akár PSII útján (Allakhverdiev et al., 2005), jelezve, hogy a fotodamérgezett PSII komplexek gyors javításához rövid idő alatt nagy mennyiségű ATP-re van szükség. Egy korábbi tanulmány azt mutatta, hogy a PSII hűtés által kiváltott fotoinhibíciója után a ciklikus elektronáramot (CEF) jelentősen stimulálták a gyenge fényviszonyok közötti helyreállítás során (Huang et al., 2010). Azt javasolták, hogy a CEF ezen stimulálása gyenge fényviszonyok mellett fokozza az ATP szintézisét a PSII gyors javításához (Huang et al., 2010). További bizonyítékokra van azonban szükség a CEF stimuláció sajátos szerepének tisztázásához gyenge fényviszonyok mellett a PSII fotoinhibíció után.
a CEF során a NADPH vagy a ferredoxin elektronjai a PSI körül forognak a plasztokinon medencébe. Ez az elektrontranszfer proton transzlokációhoz kapcsolódik, és protongradienst hoz létre a tilakoid membránok között (Xhamph) (Shikanai and Yamamoto, 2017). Amellett, hogy hozzájárul az ATP szintézis, egy másik funkciója a Simph a fotoszintetikus elektrontranszport lefelé szabályozása a tilakoid lumen megsavanyításával (Shikanai, 2014, 2016). Ez a szabályozás két különböző mechanizmust foglal magában: az egyik a hőenergia-eloszláshoz kapcsolódik, és a felesleges elnyelt fényenergiát a PSII antennák hőjeként eloszlatja (Takahashi et al., 2009), a másik pedig a Cyt B6/f komplex aktivitásának down-szabályozása, és szabályozza a PSI-be történő elektrontranszfer sebességét (Suorsa et al., 2012, 2016; Tikkanen és Aro, 2014). Ban ben zárvatermők beleértve Arabidopsis thaliana, a PSI ciklikus elektrontranszport két útja működik (Shikanai, 2007). Az első CEF út a pgr5-/PGRL1-függő út, amely érzékeny az antimicin a-ra (Munekage et al., 2002; Sugimoto et al., 2013), a másik pedig a kloroplaszt NADH dehidrogenáz-szerű (NDH) komplextől függ (Burrows et al., 1998; Shikanai et al., 1998). A pgr5-/PGRL1-függő út hozzájárulása jelentősebb a C3 növényekben. Nagy fényviszonyok mellett a PGR5-/PGRL1-függő CEF aktiválása indukálja a tilakoid lumen savasodását, és így a P700 oxidációs arány magas szintjéhez vezet (Suorsa et al., 2012, 2016; Kono et al., 2014; Yamori et al., 2016). Összehasonlításképpen, a P700 oxidációs arányt gyenge fényviszonyok mellett kevéssé befolyásolta a pgr5-/PGRL1-függő CEF hiánya (Munekage et al., 2002, 2004; Kono et al., 2014). Mérsékelt PSII fotoinhibíció esetén a CEF stimulálása gyenge fényviszonyok mellett a P700 oxidációs arány növekedésével járt (Huang et al., 2010). Nem világos azonban, hogy ezt a megnövekedett P700 oxidációs arányt a CEF stimuláció okozza-e.
a CEF mellett az ELEKTRONTRANSZFER a PSII-ből (ETRII) fontos szerepet játszik a PSI redox állapotának befolyásolásában (Tikkanen et al., 2014; Huang et al., 2016A, c; Suorsa et al., 2016). Az A. thaliana pgr5 növényeiben a PSII súlyos fotoinhibíciója valószínűleg a fotoszintetikus elektrontranszfer végső kontrolljaként működik, lehetővé téve a P700 fenntartását optimálisan oxidálva túlzott fény alatt (Tikkanen et al., 2014). Nemrég, Suorsa et al. (2016) jelezte, hogy a pgr5 növények magas P700 oxidációs arányt mutattak nagy fény mellett, amikor az ETRII-t csökkentették a psbo1, PsbP2, PsbQ1, PsbQ2 és PsbR lókuszok kiütéses mutációinak kombinálásával. A dohány hűtött leveleiben a mérsékelt PSII fotoinhibíció csökkentette az elektron áramlását a PSI-be, majd növelte a P700 oxidációs arányt a további hűtési kezelések során (Huang et al., 2016A). Ezenkívül a PSII hűtés által kiváltott fotoinhibíciója az ETRII depressziójához vezetett, amelyet a P700 oxidációs arány növekedése kísért gyenge fényviszonyok mellett (Huang et al., 2010). Feltételezzük, hogy a P700 oxidációs arányt gyenge fényviszonyok mellett elsősorban a PSII elektronáramlása szabályozza.
korábbi jelentésünkben megfigyeltük, hogy a Panax notoginseng árnyékot létrehozó növény szelektív fotoinhibíciót mutatott a PSII-ben nagy fényterhelés mellett. Rövid távú nagy fényterhelésnek való kitettség után az etrii gyenge fényviszonyok mellett jelentősen csökkent, de a CEF jelentősen stimulálódott. Ezzel párhuzamosan a P700 oxidációs arány nagymértékben megnőtt. Konkrét célunk az volt, hogy (1) megvizsgáljuk, hogy a CEF stimuláció gyenge fényviszonyok között elsősorban megkönnyíti-e az ATP szintézisét; (2) határozza meg, hogy a P700 oxidációs arány növekedése mérsékelt PSII fotoinhibíció esetén inkább az ETRII-hez kapcsolódik, mint a CEF stimulációhoz vagy a pmf változásához. E kérdések megválaszolása érdekében a Panax notoginseng ép leveleit 2258-on kezeltük 6mol foton m – 2 s – 1 30 percig. A magas fényű kezelés előtt és után meghatároztuk az energiaeloszlást a PSII-ben, a PSI redox állapotát és a protonmotív erőt (PMF) gyenge fényviszonyok mellett, 54 MHz-es m-2 s-1 foton mellett.
anyagok és módszerek
növényi anyagok és Növekedési állapot
a jelen vizsgálatban 2 éves növények egy árnyék-létrehozó növény Panax notoginseng (Burkill) F. H. Chen ex C. Chow és W. G. Huang használtunk kísérletek. Ezeket a növényeket 10% – os napfényes fényviszonyok mellett termesztették (a napközbeni maximális fényintenzitás körülbelül 200 km volt) fotonok m-2 s-1). Ezekben a növényekben nem tapasztaltak víz-és táplálkozási stresszt. A fotoszintetikus mérésekhez 9 hetes, teljesen kitágult leveleket használtunk.
PSI és PSII mérések
Az I. Fotorendszer és a PSII paramétereit 25 CAC-on mértük kettős Pam-100 mérőrendszerrel (Heinz Walz GmbH, Effeltrich, Németország). A klorofill fluoreszcencia paramétereit a következőképpen számítottuk ki: Fv / Fm = (Fm-Fo)/Fm, Y(II) = (Fm’ – Fs) / Fm’ (Genty et al., 1989), NPQ = (Fm – Fm’)/Fm’. Az FO és az Fm a minimális és a maximális fluoreszcencia a sötét akklimatizáció után. Fs a fényhez igazított egyensúlyi állapotú fluoreszcencia. Az FO-t és az Fm-t sötét akklimatizáció után határoztuk meg 30 percig a magas fényű kezelés előtt és után. A PSI fotoszintetikus paramétereit kettős PAM-100-mal mértük P700 jel alapján (830 és 875 nm-es impulzusmodulált fényerősség-különbség a fotodetektort elérve) (Klughammer and Schreiber, 2008). A P700 + jelek (P) a minimális (P700 teljesen redukált) és a maximális (P700 teljesen oxidált) szint között változhatnak. A maximális szintet (Pm) telítési impulzus alkalmazásával határoztuk meg (600 ms és 10000 MHz foton m-2 s-1) távoli vörös fénnyel történő elővilágítás után, és Pm-t használtunk a PSI aktivitás becsléséhez. A Pm ‘ -et a Pm-hez hasonlóan határoztuk meg, de a távoli vörös fény helyett aktin fénnyel. A PSI kvantumhozamát Y (I) = (Pm’- P)/Pm-ként számítottuk ki. A P700 oxidációs arányt egy adott aktin fényben Y (ND) = P/Pm-ként számítottuk ki. Az akceptoroldali korlátozás miatti psi nem fotokémiai energiaeloszlás kvantumhozamát Y (NA) = (Pm – Pm’)/Pm-ként számítottuk ki. Az Y(II), NPQ, Y(I) és Y(ND) egyensúlyi állapot értékeit gyenge fényviszonyok között 20 percig tartó fényakklimatizáció után mértük.
Fotoszintetikus elektron áramlását PSZI és PSII volt számítása a következőképpen történik: ETRII = Y (II) ×PPFD ×0.84 ×0.5 (diana Krall, valamint Edwards, 1992), ETRI = Y (I) ×PPFD×0.84 ×0.5 (Yamori et al., 2011), ahol feltételezzük, hogy 0,5 a PSI-t vagy PSII-t elérő elnyelt fény aránya, 0,84 pedig az abszorpció (a levelek által elnyelt beeső fény frakciója). A CEF aktiválásának mértékét etri/ETRII arányban becsülték meg (Yamori et al., 2011, 2015). Meg kell jegyezni, hogy a PSI és a PSII közötti 1:1 gerjesztő particionálás nem valós, mert az árnyékoló növények általában növelik a PSII/PSI arányt (Lunde et al., 2003; Tikkanen et al., 2006; Grieco et al., 2012). Továbbá ebben a vizsgálatban a PSI és PSII paramétereket a Dual-Pam 100 (Walz, Németország) által biztosított vörös aktin fénnyel (635 nm) mértük. Mivel a vörös aktin fény kedvez a PSII gerjesztésének a PSI-vel szemben, a gerjesztési energia eloszlását az LHCII-től a PSII-ig és a PSI-ig befolyásolhatja a vörös aktin fény (Tikkanen et al., 2017).
Elektrokróm Shift analízis
az ECS jelet 515 nm-es abszorbancia-változásként figyelték meg egy P515/535 emitter-detektor modullal (Walz) felszerelt Dual-PAM-100 (Walz, Effeltrich, Németország) alkalmazásával. Az ECS jelet 20 perc megvilágítás után kaptuk 54 m-2 s-1 fotonon, utána az ECS bomlását az aktinikus fény kikapcsolásával mértük 30 s.az ECS sötét intervallum relaxációs kinetikájának (DIRKECS) elemzését Sacksteder módszerével végeztük et al. (2001) és Takizawa et al. (2008). A teljes pmf-et az ECS jel gyors bomlásának teljes amplitúdójából becsülték 300 ms sötét intervallum alatt. Az ECS jel lassú relaxációja lehetővé tette a proton gradiens hozzájárulásának felismerését a tilakoid membránokon keresztül (Xhamph). Az elsőrendű ECS relaxáció (tECS) időállandója fordítottan arányos a tilakoid membrán proton vezetőképességével (gH+) az ATP-szintázon keresztül (Sacksteder and Kramer, 2000; Cruz et al., 2005). Ennek eredményeként gH+ a bomlási idő Állandó inverzének becsülték .
Fotoinhibitor kezelések
30 percig tartó sötét akklimatizáció után az FV / Fm és Pm értékeket ép levelekben mértük. Ezt követően ezeket az ép leveleket 59 MHz-es m-2 s-1 fotonon 20 percen át fényhez igazítottuk, és a klorofill fluoreszcencia, a P700 jel és az ECS jel paramétereit rögzítettük. Ezután az aktinikus fényt 2258 db M-2 s-1 fotonra változtattuk. Miután ezt a nagy fényt 30 percig kitettük, az aktinikus fényt azonnal 59 m-2 s-1-re változtattuk, és a fotoszintetikus paramétereket 20 percig tartó fény akklimatizáció után rögzítettük. Végül az Fv/Fm-et és a Pm-et 30 percig tartó sötét akklimatizáció után mértük.
statisztikai elemzés
az eredményeket öt független kísérlet átlagértékeként jelenítettük meg. Független t-tesztet alkalmaztunk 6,05 szignifikancia szinten annak meghatározására, hogy jelentős különbségek voltak-e a különböző kezelések között. Az összes statisztikai elemzést SPSS 16.0 alkalmazásával végeztük.
eredmények
a fotoszintetikus indukció során 59 dB m-2 s-1 fotonon az ETRII fokozatosan emelkedett, és körülbelül 18 perc elteltével elérte az egyensúlyi állapotot (1a.ábra). Összehasonlításképpen, az etri állandó volt ebben az indukciós fázisban (1a ábra). A gyenge fény megjelenése után az ETRI sokkal magasabb volt, mint az ETRII. 20 perces fotoszintetikus indukció után azonban az ETRI alacsonyabb volt, mint az ETRII. Ezek az eredmények a CEF aktiválását javasolták az indukció kezdeti szakaszában, a korábbi vizsgálatokkal összhangban (Joliot and Joliot, 2002, 2005; Makino et al., 2002). Ezt követően fotoszintetikus indukció gyenge fényviszonyok mellett, a leveleket 2258 6mol foton m-2 s-1 nagy fényben világítottuk meg 30 percig. Érdekes módon az ETRII fokozatosan csökkent a magas fényű kezelés során (1b ábra). Eközben az etri értéke magasabb volt, mint az ETRIIÉ (1b ábra), jelezve a CEF aktiválódását nagy fény mellett. Ezt a nagy fényű kezelést követően a maximális fotooxidálható P700 (Pm) stabil maradt (2a ábra). Összehasonlításképpen, a maximális fluoreszcencia intenzitás (Fm) 40% – kal csökkent,a PSII (Fv/Fm) maximális kvantumhozama pedig 0,80-ról 0,65-re csökkent (2b, C ábra). Ezek az eredmények jelezték a PSII szelektív fotoinhibícióját a Panax notoginseng leveleiben nagy fényterhelés alatt.
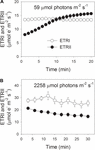
1. ábra. Fotoszintetikus elektron áramlás gyenge fényviszonyok mellett (a) és magas fényviszonyok mellett (B). (A) A Sötét akklimatizált leveleket 59 6mol foton m-2 s-1 gyenge fénynek tesszük ki 20 percig. (B) megvilágítása után 59 6mol fotonok m-2 s-1 20 percig, hogy aktiválja az elektron mosogató fotoszintézis, levelek voltak kitéve nagy fény 2258 m-2 2 m-1 30 percig. Az értékek azt jelentik, hogy (n = 5).
a nagy fényerejű kezelés előtt az érett leveleket gyenge fényviszonyok között 59 dB m-2 s-1 fotonnal világítottuk meg 20 percig a fotoszintézis aktiválásához, majd az ETRI, ETRII, Y(ND) és NPQ értékeket rögzítettük. 30 perces magas fényű kezelés után az etri, az ETRII, az Y(ND) és az NPQ értékeit egy új, 20 perces akklimatizációs fázist követően rögzítettük gyenge fényviszonyok között. A nagy fényerejű kezelés előtt az ETRI és az ETRII értéke 59 km-es m-2 s-1 fotonnál 13,4, illetve 15,7 km-es m-2 s-1 elektron volt (3a., B. ábra). Erős fényű kezelés után az ETRI és az etrii gyenge fényviszonyok mellett 12,8, illetve 11,2 db M-2 s-1 elektron volt (3a., B. ábra). Az etrii értéke 59 dB m-2 s-1 fotonnál körülbelül 30% – kal csökkent a fotoinhibitor kezelés után, jelezve az etrii depresszióját mérsékelt PSII fotoinhibíció esetén. A nagy fényerejű kezelés előtt az ETRI/ETRII arány értéke 59 dB m-2 s-1 fotonnál 0,85 volt (3C ábra). Fotoinhibitor kezelés után az ETRI/ETRII Arány szignifikánsan 1,14-re nőtt (3C ábra). Ezek az eredmények azt sugallták, hogy a CEF alacsony fényviszonyok mellett stimulálódik a PSII fotoinhibíción (Yamori et al., 2011, 2015).
a nagy fényerejű kezelés után az Y(I) egyensúlyi értéke 59 m-2 s-1 fotonnál nem változott (4a. ábra). Érdekes módon az Y (NA) szignifikánsan csökkent 0,36-ról 0,23-ra (4b ábra), az Y(ND) pedig szignifikánsan nőtt 0,1-ről 0,25-re (4c ábra). Ezek az eredmények a PSI redox állapotának változását jelezték gyenge fényviszonyok mellett fotoinhibitor kezelés után. Eközben az NPQ értéke kissé megváltozott (4D ábra). Ennek eredményeként a mérsékelt PSII fotoinhibíció eltérő hatással volt a P700 oxidációs arányára és az NPQ-ra gyenge fényviszonyok mellett.
annak tisztázása érdekében, hogy az Y(ND) növekedését a pmf növekedése okozza-e, az elektrokróm eltolódási jeleket 54 db M-2 s-1 fotonnál a fény akklimatizálása után 20 percig határoztuk meg a nagy fényű kezelés előtt és után. Érdekes módon a teljes pmf szignifikánsan 18%-kal csökkent a magas fénykezelést követően (5a.ábra), de az anti-pH szint jelentéktelen mértékben változott (5b. ábra). Mivel a pmf képződését befolyásolhatja a tilakoid proton vezetőképessége, a tilakoid membrán proton vezetőképességét (gH+) ezen gyenge fényviszonyok mellett is megvizsgáltuk a magas fényű kezelés előtt és után. Az eredmény azt mutatta, hogy a GH+ nem változott a nagy fényű kezeléssel (5c ábra), ami arra utal, hogy a nagy fényű kezelés alig befolyásolta a kloroplaszt ATP-szintáz aktivitását gyenge fényviszonyok mellett. Mivel az ETRII 30% – os csökkenését a pmf 18% – os csökkenése kísérte, a CEF gyenge fényviszonyok mellett történő stimulálása részben kompenzálta a PMF és az ONS kialakulását.

5. ábra. A teljes protonmotív erő (PMF) (a), A protongradiens (6PH) a tilakoid membránokon (B), valamint a tilakoid membrán (C) protonvezetőképessége (gH+) gyenge fényviszonyok mellett. Az összes paramétert az alkalmazkodás után mértük 54 m-2 s-1 foton m-20 perc alatt. Az értékek azt jelentik, hogy (n = 5). A csillag jelentős (p < 0,05) változást jelez nagy fényű kezelés után.
Vita
a CEF stimuláció szerepe gyenge fényviszonyok mellett
kimutatták, hogy a CEF fontos szerepet játszik a fotoszintézis és a növények növekedésének fenntartásában (Yamori and Shikanai, 2016). Nagy fényviszonyok mellett a CEF-ről úgy gondolják, hogy elengedhetetlen az ATP/NADPH energia költségvetésének kiegyensúlyozásához, valamint a PSI és a PSII károsodástól való védelméhez (Munekage et al., 2002, 2004; Takahashi et al., 2009; Suorsa et al., 2012, 2016; Walker et al., 2014; Huang et al., 2015a, 2017b). Alacsony fényintenzitás mellett a CEF fontos szerepet játszik a fotoszintetikus CO2-asszimiláció optimalizálásában, valószínűleg extra ATP-ellátás révén (Yamori et al., 2011, 2015; Nishikawa et al., 2012; Huang et al., 2015a). A CEF fő szerepe rugalmasan modulálódik a fényintenzitásra reagálva. Különösen a CEF elsősorban hozzájárul az ATP / NADPH energia költségvetésének kiegyensúlyozásához szubszaturáló fényintenzitások mellett, de főleg védi fotoszintetikus készülék fotoinhibícióval szemben, ha telítő fényintenzitásnak van kitéve a tilakoid lumen megsavanyításával (Huang et al., 2015a). Érdekes módon, Huang et al. (2010) megállapította, hogy a CEF szignifikánsan stimulálódott gyenge fényviszonyok mellett a PSII hűtés által kiváltott fotoinhibíciója után, és feltételezte, hogy ez a CEF stimuláció elsősorban fokozta az ATP szintézisét a PSII gyors javítása érdekében. Ennek a hipotézisnek a alátámasztásához azonban további bizonyítékokra volt szükség. Jelen tanulmányban megfigyeltük, hogy az etri/ETRII értéke gyenge fényviszonyok mellett jelentősen megnőtt (3C ábra), jelezve a CEF stimulációját a PSI körül (Huang et al., 2011, 2012, 2017A,b; Yamori et al., 2011, 2015). Ezzel párhuzamosan a pmf amplitúdója szignifikánsan csökkent, az Xhamph pedig enyhén csökkent. Ezek az eredmények azt mutatták, hogy a CEF stimulálása nem váltotta ki a fotoszintetikus elektrontranszportban a Naplószámtól függő Down-szabályozást. Más szavakkal, ez a CEF stimulálása gyenge fényviszonyok mellett elsősorban megkönnyítette az ATP szintézisét.
bár a kloroplaszt ATP-szintáz protonvezetőképességét a nagy fényű kezelés nem befolyásolta, a pmf kisebb mérete az ATP szintézis tényleges sebességének csökkenését eredményezte. Az árak a PSII javítás és fotoszintetikus CO2 asszimiláció gyenge fényviszonyok között elsősorban korlátozza a sebesség ATP szintézis (Allakhverdiev et al., 2005; Yamori et al., 2011, 2015; Nishikawa et al., 2012). Mivel az ETRII jelentősen csökkent a PSII fotoinhibíció esetén, az ETRII-n keresztüli ATP szintézis sebessége jelentősen csökkent. Az elsődleges anyagcseréhez és a PSII gyors helyreállításához szükséges ATP/NADPH Arány kiegyensúlyozásához rugalmas mechanizmusra van szükség az extra ATP biztosításához. Ebben az állapotban az ETRII 30% – kal csökkent, de a teljes pmf csak 18% – kal csökkent. Ezek az eredmények azt mutatják, hogy a CEF stimulálása kompenzálta a PMF etrii-függő képződésének csökkenését, fokozva az ATP szintézisét. A fotodamérgezett PSII gyors javítása az ATP szintézisétől függ (Allakhverdiev et al., 2005). A CEF gyorsulása a PSI körül növelte az ATP intracelluláris koncentrációját, ezáltal felgyorsítva a Psii javulásának sebességét a Synechocystis-ban (Allakhverdiev et al., 2005). Összességében azt javasoljuk, hogy a CEF stimulálása gyenge fényviszonyok mellett fontos szerepet játszik a PSII aktivitás gyors javításában egy további ATP szintézis révén.
A P700 oxidációs arányának változása gyenge fényviszonyok mellett nem kapcsolódik a pmf-hez
érdekes jelenség, hogy a P700 oxidációs Arány gyenge fényviszonyok mellett jelentősen megnőtt mérsékelt PSII fotoinhibíció esetén (Huang et al., 2010), amelyet a jelen tanulmány is mutat. Olyan körülmények között, amikor az elnyelt fény meghaladja a fotoszintézis követelményeit, a CEF-függő generálása a CET B6/f komplex aktivitását szabályozza, és szabályozza az elektron áramlását a PSII-től a PSI-ig (Suorsa et al., 2012, 2016; Shikanai, 2014, 2016), optimalizálja a P700 redox állapotát a PSI-ben és minimalizálja a PSI ros termelését a fotoszintézis során. Az A. thaliana pgr5-növényeiben a P700-at elektronok csökkentik a fényben a PMF-generáció elvesztése miatt. Azonban a pgr5 mutáns felhalmozódó flavodiiron proteinben a PMF szintje szintén visszaállt a vad típusú szintre, így a P700 oxidálódott, mint a vad típusú (Yamamoto et al., 2016). Ban ben klorofill b-hiányos búza mutáns vonalak, az elégtelen tilakoid proton gradiens a PSI akceptor oldal túlzott redukciójához vezet, így psi fotoinhibíció nagy fény vagy magas hőmérséklet mellett (Brestic et al., 2015, 2016). Nemrégiben néhány tanulmány arról számolt be, hogy a kloroplaszt ATP-szintáz fokozott aktivitása károsította a pmf képződését, és a fotoszintetikus elektrontranszport lánc túlzott redukcióját okozta, ami a PSI fotodamázsát eredményezte nagy fény és ingadozó fény alatt (Kanazawa et al., 2017; Takagi et al., 2017). Ezek a jelentések alátámasztják a PMF kritikus szerepét a P700 optimális oxidációjának fenntartásában a felesleges fényenergia alatt.
jelenlegi eredményeink azt mutatták, hogy a 30 percig tartó magas fényű kezelés után a gyenge fényviszonyok mellett képződött teljes pmf jelentősen csökkent, és az ONS kialakulása kissé megváltozott (5A., B. ábra). Eközben a P700 oxidációs Arány szignifikánsan nőtt 0,1-ről (kezelés előtt) 0,25-re (kezelés után) (4c ábra). Ezek az eredmények határozottan azt mutatják, hogy a P700 oxidációs arányának növekedése nem magyarázható a pmf változásával, amely nagymértékben különbözik az Y(ND) és a PMF közötti korrelációtól nagy fényviszonyok mellett (Yamamoto et al., 2016; Takagi et al., 2017). Ezért a PMF szabályozó hatása a P700 oxidációs arányra gyenge fényviszonyok mellett minimális, de nagy fény vagy ingadozó fény esetén különösen fontossá válik. Magas fény mellett az Y(ND) magas értékét általában az NPQ magas értéke kíséri (Munekage et al., 2002, 2004; Kono et al., 2014; Zivcak et al., 2015), a pmf növekedése miatt. Az a tény, hogy a képződés csökkenése miatt csökken az Y(ND) és az NPQ szintje nagy fényviszonyok mellett (Suorsa et al., 2012; Kono et al., 2014; Yamamoto et al., 2016; Kanazawa et al., 2017). Az NPQ indukcióját gyenge fényviszonyok mellett elsősorban a lumen savasodásának szintje határozza meg. Nagy fényerejű kezelés után a lumen savasodásának mértéke nem változott (5b.ábra), az NPQ stabil maradt (4D. ábra). Összehasonlításképpen, Y (ND) jelentősen nőtt. Ezek az eredmények azt mutatják, hogy gyenge fényviszonyok mellett az Y(ND) és az NPQ-t különböző szabályozási mechanizmusok szabályozták.
az ETRII szabályozza a P700 oxidációs arányt gyenge fényviszonyok mellett
a PSI csak akkor csökken túlzottan, ha a PSII-ből származó elektronáramlás meghaladja a PSI elektron akceptorok képességét az elektronokkal való megbirkózáshoz. Ha az elektron áramlása a PSI-hez korlátozott, a PSI rendkívül toleráns a könnyű stressz ellen az A. thaliana pgr5 növényeiben (Suorsa et al., 2012, 2016; Tikkanen et al., 2014). Az árnyékot létrehozó növény számára Pszichotria rubra, a PSI aktivitása nem volt érzékeny a nagy fényfeszültségre DCMU jelenlétében (Huang et al., 2016c). A dohány hűtött leveleiben a mérsékelt PSII fotoinhibíció lehetővé tette a P700 optimálisan oxidált, majd védett PSI aktivitásának fenntartását a további fotokárosodás ellen (Huang et al., 2016A). Ezek az eredmények azt mutatták, hogy az ETRII fontos szerepet játszik a PSI redox állapotának szabályozásában nagy fény vagy hűtés-fény feszültségek mellett. Jelenlegi eredményeink azt mutatták, hogy a mérsékelt PSII fotoinhibíció az etrii jelentős depressziójához vezetett gyenge fényviszonyok mellett. Eközben a PSI aktivitás és a kloroplaszt ATP-szintáz aktivitása kissé megváltozott, és a teljes PMF és a Xhamsterph nem növekedett. Ennek eredményeként az etrii csökkenését nem a CYT B6/f komplexen keresztül történő Monoph-függő fotoszintetikus kontroll okozta, hanem valószínűleg a PSII aktivitás csökkenése okozta (Tikkanen et al., 2014). A PSII-től a PSI-ig terjedő elektronellátás csökkenése a P700 oxidáció fokozott szintjéhez vezetett. Így a P700 oxidációs arányt gyenge fényviszonyok mellett nagyrészt az ETRII szabályozta.
következtetés
megállapítottuk, hogy a PSII szelektív fotoinhibíciója a CEF stimulációját és a P700 oxidációs arány növekedését indukálta gyenge fényviszonyok mellett. A CEF stimulálása nem váltotta ki a fotoszintetikus elektrontranszportban az anti-pH-függő down-szabályozást. Ennek eredményeként a CEF ezen stimulálása gyenge fényviszonyok mellett elsősorban megkönnyítette az ATP szintézisét, ami elengedhetetlen a fotodamérgezett PSII gyors javításához. A P700 oxidációs arányának növekedése nem magyarázható a CYT B6/F komplexben a CA-tól függő fotoszintetikus kontroll változásával, hanem elsősorban a PSII-től a PSI-ig terjedő elektronellátás csökkenése okozta.
szerzői hozzájárulások
WH, S-BZ és TL fogant és tervezett kutatás. WH és Y-JY végezte a kísérleteket. WH, Y-JY, S-BZ és TL elemezte az adatokat. WH írta a kézirat első tervezetét, amelyet az összes szerző intenzíven szerkesztett.
finanszírozás
ezt a tanulmányt a kínai Nemzeti Természettudományi Alapítvány (31670343), a Kínai Tudományos Akadémia Ifjúsági Innovációs Promóciós Szövetsége (2016347), valamint a Yunnan tartomány főbb tudományos és technikai programjai (2016zf001) támogatták.
összeférhetetlenségi nyilatkozat
a szerzők kijelentik, hogy a kutatást olyan kereskedelmi vagy pénzügyi kapcsolatok hiányában végezték, amelyek potenciális összeférhetetlenségnek tekinthetők.
Kono, M., Noguchi, K., and Terashima, I. (2014). A ciklikus elektron áramlás szerepe a PSI (CEF-PSI) és az O2-függő alternatív utak körül a fotoszintetikus elektron áramlás szabályozásában rövid távú ingadozó fényben az Arabidopsis thaliana-ban. Növényi Sejt Fiziol. 55, 990–1004. doi: 10.1093 / pcp / pcu033
PubMed absztrakt / CrossRef teljes szöveg / Google Tudós
Shikanai, T. (2016). A protonmotívum szabályozó hálózata: a ciklikus elektrontranszport hozzájárulása a fotorendszer körül I. Photosynth. Res. 129, 253-260. doi: 10.1007 / s11120-016-0227-0
PubMed Abstract | CrossRef Full Text / Google Scholar
Yamori, W., Makino, A., and Shikanai, T. (2016). Az I. fotorendszer körüli ciklikus elektrontranszport fiziológiai szerepe a fotoszintézis fenntartásában a rizs ingadozó fénye alatt. Sci. Ismétlés 6:20147. doi: 10.1038 / srep20147
PubMed absztrakt | CrossRef teljes szöveg | Google Tudós
Yamori, W., and Shikanai, T. (2016). Az I. fotorendszer körüli ciklikus elektrontranszport élettani funkciói a fotoszintézis és a növénynövekedés fenntartásában. Annu. Rev. Növény Biol. 67, 81–106. doi: 10.1146 / annurev-arplant-043015-112002
PubMed absztrakt | CrossRef teljes szöveg | Google Tudós
Yamori, W., Shikanai, T., and Makino, A. (2015). Az I. fotorendszer ciklikus elektron áramlása kloroplaszt NADH dehidrogenáz-szerű komplexen keresztül fiziológiai szerepet játszik a fotoszintézisben gyenge fényviszonyok mellett. Sci. REP. 5:13908. doi: 10.1038 / srep13908
PubMed absztrakt | CrossRef teljes szöveg | Google Tudós
Zhang, S. P., and Scheller, H. V. (2004). A fotorendszer fotoinhibíciója lehűlési hőmérsékleten, majd az azt követő helyreállítás az Arabidopsis thaliana-ban. Növényi Sejt Fiziol. 45, 1595–1602. doi: 10.1093/pcp/pch180
PubMed absztrakt | CrossRef teljes szöveg | Google Tudós