- Introduksjon
- Metode
- Deltakere
- Vurdering Av Kreativitet
- Torrance Test Av Kreativ Tenkning (Sirkler Underskala)
- Alternativ Bruk Oppgave (AUT)
- Pretest
- dna-ekstraksjon og genotyping
- Statistisk Analyse
- Resultater
- Flyt Komponent
- Fleksibilitetskomponenten
- Originalitetskomponenten
- Kjønnsforskjeller
- Diskusjon
- Interessekonflikt
Introduksjon
Kreativ kognisjon spiller en viktig rolle i kunsten, i oppfinnelse Og innovasjon, så vel som i hverdagen (Runco Og Richards, 1997). Selv om kreativitet har blitt ansett som en unik menneskelig kapasitet, spontant kreative atferd (f. eks., å skape nye verktøy blant primater og fugler) har vist seg å forekomme hos ikke-mennesker også (Byrne og Bates, 2007), som videre bekrefter kreativitetens dype evolusjonære og biologiske røtter. Kreativitet er definert som evnen til å produsere svar som er både nye (dvs. originale, sjeldne og uventede) og passende(dvs. adaptive og nyttige i henhold til oppgavebegrensninger) (Sternberg og Lubart, 1999). En av de psykometriske tilnærmingene til måling av kreativitet innebærer divergerende tenkning (DT) oppgaver der deltakerne blir bedt om å svare på et gitt problem med flere løsninger (Dietrich og Kanso, 2010). Tester av DT måler generelt ulike aspekter av kreativitet, inkludert kreativ flyt, fleksibilitet og originalitet (Torrance, 1974). SÅLEDES, selv om DET ikke er synonymt med kreativitet, gir dt-oppgaver strukturerte og objektive målinger av kreativitet (Sternberg Og Lubart, 1999; Jung et al., 2009). Spesielt har score PÅ dt-oppgaver vist seg å være positivt korrelert med økologisk gyldige tiltak for kreativ prestasjon (Carson et al., 2005) så vel som med egenverdig kreativitet(Furnham og Bachtiar, 2008).
Tidligere studier har pekt på involvering av dopaminergic (DA) system i kreativitet (Heilman et al., 2003; Flaherty, 2005; Takeuchi et al., 2010). Takeuchi et al. (2010) fant individuelle forskjeller i kreativitet, målt VED DT, å være positivt korrelert med grå materie I DA-systemregioner, inkludert den dorsolaterale prefrontale cortex, bilaterale basale ganglia, substantia nigra og tegmental ventralområdet. I tillegg har flere genetiske studier vist et forhold MELLOM dt og dopamin nevrotransmisjon (Reuter et al., 2006; De Manzano et al., 2010; Runco et al., 2011). For eksempel, Reuter et al. (2006) fant kreativitet, målt ved DT-oppgaver som involverer både figural og verbal kreativitet, å være signifikant forbundet med polymorfier av dopamin D2-reseptorgenet (DRD2). Videre har flere studier funnet kortikal dopamin å være involvert i kognitiv fleksibilitet (Frank, 2005; Cools, 2008; Garcia-Garcia Et al., 2010), en av hovedkomponentene I DT.Et interessant DA-kandidatgen for kreativitet er dopamin D4-reseptorgenet (DRD4). DRD4-reseptoren er en av fem dopaminreseptorer og spiller en viktig rolle i å formidle synaptisk dopamin-signalering. Genet er preget av et 48 basepar variabelt antall tandemrepetisjoner (48-bp VNTR) lokalisert i kodingsområdet til den tredje eksonen. DRD4 48-bp vntr polymorfisme varierer fra 2 til 11 gjentar på tvers av individer (Asghari et al., 1995; Cravchik Og Goldman, 2000), hvor 4-repeat (4R) er den vanligste gjentakelsen i Kaukasiske befolkninger, og 7-repeat (7R) er den nest vanligste varianten (Chang et al., 1996; Ding et al., 2002). Interessant NOK HAR 7R tidligere vært assosiert med virkelige kreative atferd, for eksempel nyhetssøkende personlighetstrekk (Ebstein et al., 1996). Faktisk er nyhetssøkende-tendensen til utforskende aktivitet-antatt å være et av egenskapene til kreative mennesker (Chavez-Eakle et al., 2006; Drago et al., 2009). Selv om bevisene for en sammenheng MELLOM DRD4 7R og nyhetssøkende er inkonsekvent, som det fremgår av flere meta-analyserapporter (Kluger et al., 2002; Munafò et al., 2008), har mange studier funnet en betydelig sammenheng mellom nyhetssøkende OG 7R-allelen TIL DRD4 (Ebstein et al., 1996; Benjamin et al., 2000; Keltikangas-J ③rvinen et al., 2002; Becker et al., 2005). Roussos et al. (2009) har nylig antydet at forskjeller i måling av nyhetssøkende ved hjelp av selvrapporteringsskalaer kan utgjøre de inkonsekvente resultatene MELLOM 7r-polymorfisme og nyhetssøkende. I tråd med dette har DET gjentatte ganger blitt funnet AT DA-systemet spiller en viktig rolle i personlighetstrekk av nyhetssøkende (Flaherty, 2005; Schweizer, 2006), som videre bekrefter DRD4S potensielle rolle i kreativitet.likevel, i motsetning TIL ROLLEN SOM DRD4 48-bp VNTR 7R i nyhetssøkende, indikerer nyere bevis AT 7R faktisk er forbundet med nedsatt kognitiv fleksibilitet, en av de viktigste aspektene av kreativitet (Strobel et al., 2004; Congdon et al., 2008). Fleksibilitet innebærer evnen til å gjøre en alternativ respons etter vellykket hemme en gjeldende respons. Funn så langt på sammenhengen mellom hemming OG DRD4 er blandet. Congdon et al. (2008) fant at deltakere MED 7R-allelet I DRD4 utviser høyere stopp-signalreaksjonstid (SSRT) på en go / no-go-oppgave, noe som reflekterer dårligere hemmende kontroll, Mens Forbes et al. (2009) Klarte ikke å finne en slik effekt på Barratt Impulsiveness Scale (BIS), et selvrapporteringsverktøy for å måle impulsivitet. På den annen side, Colzato et al. (2010) viste at disse blandede effektene kan skyldes at tidligere studier adresserte impulsivitet som en monolitisk prosess, mens impulsivitet faktisk kan deles inn i funksjonelle og dysfunksjonelle typer i Henhold Til Dickmans Impulsivitetsbeholdning (DII; Dickman, 1990). Dysfunksjonell impulsivitet er tendensen til å handle uten omtanke i tilfeller der slik handling er upassende, mens funksjonell impulsivitet er en lignende tendens implementert i passende situasjoner (Colzato et al., 2010). Colzato et al. (2010) fant at personer MED DRD4 7R, som er forbundet med høyere nivåer av striatal DA, viste høyere dysfunksjonell impulsivitet. Flere meta-analytiske studier har pekt PÅ 7R allelet AV DRD4 som en risiko allel for attention-deficit hyperactivity disorder (ADHD; Faraone et al., 2001; DiMaio et al., 2003), en lidelse preget av høy dysfunksjonell impulsivitet (Young et al., 2007). Videre har forholdet MELLOM ADHD og kreativitet blitt undersøkt i flere studier (Shaw And Brown, 1990; Healey og Rucklidge, 2006), men så langt har disse studiene gitt uklare resultater. MENS NOEN har funnet ADHD og dets symptomologi å være positivt korrelert med kreativitet (Healey og Rucklidge, 2006; White og Shah, 2011), har andre rapportert en motsatt trend (Funk et al.(1993; Healey og Rucklidge, 2005, 2008). INTERESSANT, ADHD er atferdsmessig forbundet med svekkelser i utøvende kontroll, inkludert attentional set-shifting (Boonstra et al., 2005), og fleksibilitet (Barkley et al., 1997; Sersjant et al., 2002), som begge er viktige for kognitiv fleksibilitet, en sentral dimensjon av kreativitet (Dietrich, 2004; Durstewitz og Seamans, 2008). Disse funnene indikerer at personer MED 7R-allelet I DRD4 kan vise lavere nivåer av kreativitet og redusert fleksibilitet på DT-oppgaver spesielt. DERFOR ER DRD4 exon III VNTR en biologisk plausibel kandidat for å bidra til individuelle forskjeller i kreativitet.Samlet sett ser DET Ut TIL AT 7R-allelet, SOM en risikoallel for svekkede eksekutive funksjoner, fleksibilitet og settforskyvning, kan være forbundet med lav kreativitet. På den annen side indikerer sammenhengen MELLOM 7r-allelet og høyere nyhetssøkende at personer med 7R-allelet faktisk kan vise større fleksibilitet og kreativitet.den nåværende studien ble designet for å utforske drd4 7RS komplekse rolle i delvis å forme menneskelig kreativitet. For å bedre karakterisere dette forholdet, vurderte vi to typer dt-tester(figural og verbal). Scoring av hver oppgave ble delt i henhold til DE tre hoveddimensjonene AV DT, nemlig kreativ flyt, fleksibilitet og originalitet, og hver dimensjon ble analysert separat for å teste hypotesen OM AT DRD4 differensielt påvirker hver dimensjon av kreativitet.
Metode
Deltakere
prøven besto av 185 studenter Fra Universitetet I Haifa I Israel (112 kvinner). Alle deltakerne Var Kaukasiske (selvrapporterte), med en gjennomsnittsalder på 24,5 (SD ± 2,1). Deltakerne ble rekruttert gjennom kunngjøringer lagt ut på universitetet. Alle deltakerne ble betalt frivillige.
Vurdering Av Kreativitet
Kreativitet ble vurdert ved hjelp av to dt-tester, en delskala fra figural sub-test Av Torrance test of creative thinking (Torrance, 1974), Og Alternativ Bruker Oppgave (Aut; Guilford et al., 1978). Begge testene inkluderte DE tre kjernedimensjonene TIL DT, nemlig fleksibilitet, originalitet og flyt.
Torrance Test Av Kreativ Tenkning (Sirkler Underskala)
Deltakerne fikk en side hvor 30 identiske sirkler ble tegnet. De ble bedt om å tegne så mange forskjellige meningsfulle objekter som mulig innen en frist på 10 min, hvor hver tegning må inneholde minst en sirkel. Scoring inkludert flyt (antall svar produsert), fleksibilitet (antall kategorier) og originalitet, beregnet i henhold til scoring av opprinnelige svar, som beskrevet i Torrance Tests Of Creative Thinking scoring guide (Torrance, 1974).
Alternativ Bruk Oppgave (AUT)
Deltakerne ble gitt en liste over fem vanlige objekter og bedt om å liste så mange alternative bruksområder som mulig for hvert objekt, innen en frist på 10 min. Den vanligste hverdagsbruken ble angitt i parentes. Objektene var: sko( vanlig bruk: slitasje til fots); knapp (vanlig bruk: lukke ting); blyant (vanlig bruk: tegning eller skriving); dekk (vanlig bruk: bilhjul); og drikkeglass (vanlig bruk: inneholder væske). Bare svar som ikke gjentok de gitte vanlige bruksområdene ble talt og inkludert. Som I Torrance-testen inkluderte scoring flyt, fleksibilitet og originalitet. Siden DET ikke er noen retningslinjer for scoring av originale svar i AUT, ble originale svar definert som statistisk sjeldne svar i henhold til en pretest utført i laboratoriet vårt.
Pretest
for å skape et gyldig kriterium for responsfrekvens, fullførte en gruppe på 100 friske deltakere som ikke deltok i denne studien AUT. For hvert objekt ble en liste over alle mulige bruksområder samlet fra alle deltakere. Et statistisk sjeldent mål ble beregnet basert på denne listen for å evaluere originality score for hvert svar, og deretter for hver deltaker. Svarene ble tildelt en score på null hvis 5% eller flere av deltakerne oppførte en gitt bruk, en score på en hvis mellom 2% og 4,99% av deltakerne oppførte den, og en score på to hvis mindre enn 1,99% oppførte bruken. En gjennomsnittlig originalitetsscore ble beregnet for hver deltaker i henhold til disse statistiske sjeldne scorene.
dna-ekstraksjon og genotyping
DNA ble ekstrahert fra 20 ml munnvannsprøver ved Hjelp Av Master Pure kit (Epicentre, Madison, WI). DRD4 48-bp VNTR ble karakterisert VED EN PCR forsterkning prosedyre med følgende primere: F5 ‘- CTT CCT ACC CTG CCC GCT CAT GCT gctgct CTA CTGG – 3 ‘Og R5’ – ACC ACCACC GGC AGG ACC CTC ATG GCC TTG CGC TC-3. PCR-reaksjonene ble utført ved bruk av 5 µ master Mix( Thermo scientific), 2 µ primere (0,5 µ), 0,6 µ mg/Cl2 (2,5 mM), 0.4 ④l DMSO 5% og 1 µ vann til totalt 9 µ totalt volum, og ytterligere 1 µ av genomisk DNA ble tilsatt til blandingen. ALLE PCR-reaksjoner ble utført på Et Biometra T1-Termosykler(Biometra, Gü, Tyskland). PCR-reaksjonstilstanden var som følger: forvarmingstrinn ved 94.0°C i 5 min, 34 sykluser med denaturering ved 94.0 hryvnias C i 30 s, reannealing ved 55 hryvnias C i 30 s, og forlengelse ved 72 hryvnias C i 90 s. reaksjonen fortsatte å holde ved 72 hryvnias C i 5 min. Reaksjonsblandingen ble deretter elektroforesert på en 3% agarosegel (AMRESCO) med etidiumbromid for å screene for genotyper.
fordelingen av genotypefrekvens var I Henhold Til Hardy-Weinberg-likevekten (chi-kvadrat = 0,01, p-verdi = 0,92).
Statistisk Analyse
Genotype ble klassifisert i henhold til nærvær eller fravær AV 7R allelet AV DRD4 (7 vs. no7), som i tråd med tidligere rapporter i litteraturen (Ding et al ., 2002). Gruppe forskjeller på kreativitet score ble analysert statistisk Ved Hjelp Av ANOVAs, med tilstedeværelse (7) eller fravær (no7) AV DRD4 7R allel som uavhengig variabel. For å undersøke de ulike aspektene AV dt (flyt, fleksibilitet, originalitet), gjennomførte vi en multivariat analyse av varians (manova) separat for hver komponent.
Resultater
for å bekrefte at de to gruppene (7, no7) ikke var signifikant forskjellige når det gjelder alder og utdanning, gjennomførte vi uavhengige t-tester for å sammenligne de to gruppene (Se Tabell 1 for middel og standardavvik). Denne analysen viste at de to gruppene ikke var forskjellige når det gjelder alder eller år med utdanning. Ikke-parametriske tester (Mann–Whitney) viste signifikante forskjeller i frekvensfordelingen for kjønn (Z = 2.11 ,p < 0.05).
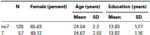
Tabell 1. Alder, kjønn og utdanningsdata FOR DRD4 7R genotype, SD: standardavvik.
middelverdien av kreativitetsmålene som brukes i de to oppgavene er oppsummert i Tabell 2.

Tabell 2. Midler Og Standardfeil (SE) av de to kreativitetsoppgavene som brukes: AUT Og Torrance.
Flyt Komponent
en multivariat ANOVA indikerte en generell effekt AV DRD4 7r genotype for flyt komponent, F(2,182) = 3,25, p < 0,05. Gitt betydningen av den samlede testen ble univariate hovedeffekter undersøkt. Signifikante univariate hovedeffekter for drd4 genotype ble oppnådd FOR AUT fluency F (1,183) = 4,95 ,p < 0.05 (7r allel viser lavere flyt i forhold til no7-gruppen, Figur 1a), Men nådde ikke nivået av betydning For Torrance fluency F(1,183) = 2.95, p = 0.087.
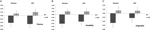
Figur 1. Kreativitet score PÅ AUT og Torrance oppgaver separert til flyt (A), fleksibilitet (B) og originalitet (C). Midler ble forvandlet Til Z-score.
Fleksibilitetskomponenten
Multivariat ANOVA av fleksibilitetstiltakene viste en generell effekt FOR drd4 7R genotype F (2,182) = 4.38 ,p < 0,05. Signifikante univariate hovedeffekter for drd4 genotype ble oppnådd FOR aut fleksibilitet F (1,183) = 4,57, p < 0,05 og For Torrance fleksibilitet F (1,183) = 6,95, p < 0,01. Som vist I Figur 1b viste i begge tilfeller individer I 7r-allelgruppen lavere fleksibilitet sammenlignet med individer i no7-gruppen (Tabell 2).
Originalitetskomponenten
Multivariat ANOVA av originalitetstiltakene viste ingen signifikanseffekter, F (2,182) = 2,06, p = 0.13, selv om resultatene indikerte lavere originalitet FOR 7R-allelet TIL DRD4 (se Figur 1c, Tabell 2 for detaljer).
Kjønnsforskjeller
Diskusjon
DT-oppgaver krever at deltakerne gir flere løsninger på et gitt problem. I oppgavene som ble brukt her, ble deltakerne pålagt å tenke på mange alternative mulige bruksområder for hverdagsartikler (I AUT), og å tegne flere tegninger som inneholder en bestemt form (sirkel) (I Torrance-oppgaven). Resultatene som presenteres her viser at personer MED DRD4 7R allelet utviser lavere kreativitet målt VED DT. Tatt I betraktning AT 7R allelet har vært assosiert med dysfunksjonell impulsivitet og dårlig inhibering (Congdon et al., 2008; Dreber et al., 2009), det faktum at personer MED 7r-allelet viser lavere kreativitet, kan forklares av deres manglende evne til å undertrykke eller hemme åpenbare (vanlige) svar, som angitt av lavere originalitetspoeng. Lavere fleksibilitet og flytende score kan indikere at når en felles respons er gitt, blir det vanskeligere å skifte til en ny kategori( som indikert av lav fleksibilitet score), noe som fører til færre svar(som indikert av lav flyt score). Ytterligere bevis for forholdet mellom kreativitet og impulsivitet kan ses i foreningen AV 7r allelet MED ADHD (Li et al., 2006), så vel som med økt impulsivitet HOS ADHD-pasienter (Langley et al., 2004). Interessant nok har kreativitet vært knyttet TIL DA-systemet i tilfelle av schizofreni (Eysenck, 1993). Selv om flere studier viser at pasienter med schizofreni og schizotyp personlighet utviser høy kreativitet (O ‘ Reilly et al., 2001; Folley and Park, 2005), nyere modeller av schizofreni og kreativitet støtter en invertert U-formet forening der kreativiteten kan være høyere med lav moderat schizotypisk egenskap, men avtar etter hvert som alvorlighetsgraden av psykopatologi i schizofreni øker (Tsakanikos Og Claridge, 2005; Stoneham and Coughtrey, 2009; Nelson and Rawlings, 2010). Faktisk, i en fersk studie Jaracz et al. (2012) rapporterte at pasienter med schizofreni viser redusert kreativitet sammenlignet med kontroller. Videre rapporterte forfatterne at nedsatt kreativitet var forbundet med lav kognitiv fleksibilitet og impulsivitet, reflektert av deres lave score På Wisconsin Card Sortering Test (WCST). Disse resultatene er i tråd med tidligere forskning som viser at pasienter med kronisk schizofreni utviser lavere kreativitet, som i tilfelle kreativ flyt ble formidlet av score på utøvende kontrolloppgaver og i tilfelle originalitet ikke var (Abraham et al., 2007). Samlet, mens mild funksjonell impulsivitet kan forbedre kreativiteten blant pasienter med schizotypiske egenskaper, spesielt ved å generere ideer som avviger fra eksemplene gitt (Abraham Og Windmann, 2008), kan den dysfunksjonelle impulsiviteten observert i alvorlig schizofreni dempe kreative evner.
resultatene av den nåværende studien støtter forholdet mellom kreativitet og varianter AV da genetiske polymorfier (Reuter et al., 2006; Runco et al., 2011). Videre indikerer resultatene i vår prøve at HOVEDVIRKNINGEN AV DRD4-7R allelet på kreativitet kan skyldes dens innvirkning på fleksibilitet. Flere studier har knyttet kognitiv fleksibilitet MED da-veier (Ashby og Isen, 1999; Dreisbach og Goschke, 2004; Mü et al., 2007). Det har blitt postulert at aktivering Av D2 – lignende reseptorer (dvs. D2, D3 og D4) reduserer Gabaergisk inhibering og letter aktivering av flere representasjoner, og dermed øker fleksibiliteten som manifestert av set-skiftende evner(Mü et al .( 2007; Seamans and Robbins, 2010). Videre Har D2 – lignende reseptorer vært knyttet til fleksibel integrering av ny informasjon (Durstewitz og Seamans, 2008). I tråd Med Dette, Reuter et al. (2006) fant en sammenheng mellom dopaminreseptoren D2 og kreativitet målt ved en sammensatt indeks, men om fleksibilitet spilte en viktig rolle i disse resultatene, kan det ikke bestemmes. I tillegg Runco et al. (2011) rapporterte forskjeller i kreativ flyt blant bærere av flere dopamingener, blant DEM DRD4. Det er imidlertid ikke klart fra deres rapport hvilken gruppe som viser større kreative evner. Det er viktig å merke seg at I motsetning til resultatene som presenteres her, Runco et al. (2011) fant ingen sammenheng mellom tiltak av fleksibilitet målt VED DT-oppgaver og DRD4. Denne inkonsekvensen kan forklares av forskjeller i populasjonene som brukes og gruppestørrelsene eller av forskjeller i allelfrekvenser innenfor den samplede populasjonen. Videre er det bevis som tyder på at sammenhengen mellom dopamin og DT kanskje ikke er lineær (Akbari Chermahini and Hommel, 2010, 2012), noe som indikerer at en interaksjon mellom FLERE DA-gener kan påvirke de individuelle resultatene av hver genetisk variabilitet i SAMMENHENG MED DT. I tillegg har flere studier pekt på AT DA-banene kan være kjønnsfølsomme (Tammimäki og Mä, 2011; Wang et al., 2012). Zhong et al. (2010), for eksempel, fant at kjønn modulert sammenhengen MELLOM drd4 exon3 polymorfisme og følelse av rettferdighet. I den nåværende studien fant vi ikke en interaksjonseffekt mellom DRD4-7R og kjønn, noe som indikerer at kjønn ikke har en modulerende effekt på sammenhengen MELLOM DRD4 og kreativitet i vårt utvalg. Videre viser resultatene som presenteres her mangel på kjønnsforskjeller i kreativitet ,og SPESIELT I DT (For gjennomgang, Se Baer and Kaufman, 2008).som konklusjon foreslår vi i den nåværende studien at sammenhengen mellom DRD4 og kreativitet hovedsakelig påvirkes av fleksibilitet. Resultatene av den nåværende studien er i tråd med tidligere resultater som peker på involvering AV DA-systemet i kreativitet og legger til den akkumulerende kroppen av kunnskap som tyder på genetiske påvirkninger i kreativitet.
det er flere begrensninger av studien som bør anerkjennes. For det første undersøkte den nåværende studien bare sammenhengen mellom en genetisk variabilitet I DA—systemet—DRD4-og DT. Imidlertid er det andre viktige polymorfier i striatal DA-systemet som kan knyttes TIL DT og kan bidra både på en frittstående måte og i gen-geninteraksjoner. En slik polymorfisme er dopamintransportøren (DAT1), som har vist seg å være assosiert med inhibering av retur (Colzato et al., 2010) så vel som med tiltak av impulsivitet(Gizer og Waldman, 2012). Videre, som nevnt ovenfor, er en annen mulighet som ikke ble utforsket her, de mulige gen – geninteraksjonseffekter på DT. Akbari Chermahini and Hommel (2010) rapporterte en ikke-lineær sammenheng mellom en markør for striatal dopamin og DT, noe som indikerer at forholdet kan være mer komplekst, slik at en polymorfisme av et gen kan påvirke DT gjennom mekling av et annet genpolymorfisme. Dermed bør fremtidige studier vurdere å undersøke samspillet mellom disse to polymorfismene i sammenheng med fleksibilitet og DT.
Interessekonflikt
forfatterne erklærer at forskningen ble utført i fravær av kommersielle eller økonomiske forhold som kan tolkes som en potensiell interessekonflikt.Akbari Chermahini, S., Og Hommel, B. (2010). (B) koblingen mellom kreativitet og dopamin: spontane øye blink priser forutsi og dissosiere divergerende og konvergent tenkning. Kognisjon 115, 458-465. doi: 10.1016 / j. kognisjon.2010.03.007
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Kjøler, R. (2008). Dopaminets rolle i motiverende og kognitiv kontroll av atferd. Hjerneforsker 14, 381-395. doi: 10.1177/1073858408317009
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Frank, M. J. (2005). Dynamisk dopaminmodulasjon i basalgangliene: en neurocomputational konto av kognitive underskudd i medisinert Og nonmedicated Parkinsonisme. J. Cogn. Neurosci. 17, 51–72. doi: 10.1162/0898929052880093
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Healey, D., Og Rucklidge, J. j. (2006). En undersøkelse av forholdet MELLOM ADHD symptomatologi, kreativitet og nevropsykologisk funksjon hos barn. Barn Neuropsykol. 12, 421–438. doi: 10.1080/09297040600806086
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Runco, Ma, Og Richards, R. (1997). Eminent Kreativitet, Hverdags Kreativitet Og Helse. Greenwich, CT: Ablex.
Torrance, E. (1974). Forskjeller er ikke underskudd. Lære. Coll. Rec. 75, 471–487.