- Vertsspesifikk Interaksjon mellom Rhizobium Og Plantepartnere
- Determinate Og Ubestemte Nodule Utvikling
- Vekst Av Symbiotiske Celler Involverer Forsterkning Av Vertsgenomet ved Endoredupliseringssykluser
- Forskjellige Skjebner Av Nitrogenfikserende Bakterier
- Vertspeptider Styrer Bakterioiddifferensiering
- NCR247: Et Eksempel på Multi-Target Host Effector
- Konklusjon
- Interessekonflikt
- Anerkjennelser
Vertsspesifikk Interaksjon mellom Rhizobium Og Plantepartnere
bakteriene som danner nitrogenfikserende symbiose med legume planter som tilhører ulike grupper av α-og β – proteobakterier kalles kollektivt rhizobia (Chen et al., 2003; MacLean et al., 2007). Mange α-proteobakterier er engasjert i langsiktige interaksjoner med høyere eukaryoter. Disse interaksjonene spenner fra overflatekolonisering gjennom fakultative symbiotiske forhold for å forplikte intracellulært patogen eller endosymbiont livsstil. De symbiotiske gener som kreves for noduleformasjon, vertscelleinfeksjon og nitrogenfiksering, er oppnådd ved lateral genoverføring som er den primære kilden til genetisk mangfold av rhizobia. Derfor kan rhizobia være nærmere relatert til patogener (Som Agrobacterium eller Brucella) enn til hverandre. Rhizobia har en tendens til å ha store genomer (opptil 10.5 Mbp) som i raskt voksende rhizobia er spredt på flere replikoner (MacLean et al., 2007). For eksempel Har Sinorhizobium meliloti, endosymbiont Av Medicago-arter, et tri-delt genom; et 3,65 Mbp kromosom og to megaplasmider, pSymA og pSymB (1,35 og 1,68 Mbp) som begge er uunnværlige og bærer flertallet av symbiotiske gener. Imidlertid inneholder Mange S. meliloti-stammer ytterligere ekstra mellomstore plasmider, og Dermed Kan s. meliloti-genomet inneholde opptil 9000 gener (Barnett et al., 2001; Capela et al., 2001). I motsetning til rhizobia har obligatoriske endosymbionter av insekter vanligvis et sterkt redusert (160-450 Kbp) genom som sikrer deres multiplikasjon og koder for noen få spesifikke biosyntetiske veier, inkludert de som tilfredsstiller vertens behov (Moran et al., 2008; Pris et al., 2011). Disse utrolig reduserte genomene forsterkes likevel kompensere det reduserte genomet med et polyploid DNA-innhold.
rhizobias plantepartnere tilhører Leguminosae / Fabaceae-familien. Nitrogenfikserende symbiose har utviklet seg i flere linjer, men ikke alle belgfrukter danner symbiose. Hittil er 12.000 nodulerte legume arter kjent, og hver har sin Egen Rhizobium partner (er). Symbiosen utløses av nitrogen sult av vertsplanten som må velge Sin Rhizobium partner fra milliarder bakterier i rhizosfæren. Dette oppnås ved sekresjon av flavonoid signalmolekyler fra roten som fungerer som kjemo-tiltrekkende midler, men viktigst som induktorer av Rhizobium nodulasjonsgener (Oldroyd, 2013). Disse genene er nødvendige for produksjon av bakterielle signalmolekyler; Nod-faktorene (NFs) som utløser noduleutviklingsprogrammet i vertsanlegget (Walker Og Downie, 2000). NFs er lipochitoligosakkaridmolekyler som bærer vertsspesifikke substitusjoner på de terminale sukkerrester og karakteristiske lipidkjeder, som gjenkjennes Av LysM-type vertsreseptorer og kreves både for noduleutvikling og bakteriell infeksjon. Interessant, den gamle symbiose av landplanter med arbuscular mycorrhizal (AM) sopp opererer med lignende lipochitoligosaccharide signal molekyler, Myc faktorer som oppfattes av lignende, men forskjellige lysm-type reseptorer Som NFs (Abdel-Lateif et al .(2012; Oldroyd, 2013). Myc-faktorene og NFs aktiverer en felles signalvei, men etter involvering av de vanlige symbiotiske gener konservert i planter, avviker veiene; en som fører til nodulasjon, den andre FOR am symbiose.
Planteinfeksjon og noduleformasjon er intrikate prosesser; Nod faktorer spiller forskjellige roller i nodule organogenesis og root hår infeksjon. Videre, foruten Nikk faktorer, ulike bakterielle overflate polysakkarider er avgjørende for effektiv infeksjon (Fraysse et al., 2003). I de fleste belgfrukter kommer rhizobia inn i verten via rothårene der ved invaginering av plasmamembranen dannes en infeksjonstråd (DEN) som inneholder multipliserende bakterier og vokser mot rotkortexen. En mindre hyppig og gammel infeksjonsmodus oppstår via sprekker på rotoverflaten av visse belgfrukter.
Determinate Og Ubestemte Nodule Utvikling
Nodule utvikling krever mitotisk reaktivering av kortikale celler som fører til nodule primordium dannelse som deretter skiller i nitrogen-feste rot knuter gir mikroaerob tilstand i den sentrale sonen for funksjon av oksygen sensitive nitrogenase enzymet i bacteroids. Avhengig av forbigående og vedvarende natur vertscelleproliferasjon, kan nodulene være enten determinate eller ubestemt type (Terpolilli et al., 2012; Kristian et al., 2013). Determinate knuter har ingen meristem og inneholder homogen populasjon av symbiotiske celler. Determinate knuter utvikler for eksempel På Phaseolus vulgaris og Lotus japonicus røtter.
Tvert imot opprettholdes den aktive celledeling i de ubestemte nodulene. En nodule meristem er tilstede i apikalområdet (sone I) som ved konstant generering av nye celler provoserer kontinuerlig vekst og langstrakt noduleform. Cellene som forlater meristemet, deler seg ikke lenger og går inn i en differensieringsfase. Smittetråden frigjør bakteriene i de submeristematiske cellene, som gradvis differensierer seg langs de 12-15 cellelagene i infeksjonssonen (sone II), noe som fører til utvikling av nitrogenfikserende symbiotiske celler I nodule zone III (Figur 1; Franssen et al ., 1992). Medicago sativa, m. truncatula, Vicia sativa og Pisum sativum er eksempler på planter som danner ubestemte knuter.
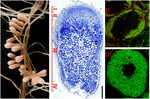
FIGUR 1. Struktur av nitrogenfikserende rotknuter dannet I s. meliloti – m. truncatula symbiose. De forskjellige nodulesonene er angitt på den langsgående noduleseksjonen: (I) meristem, (II) infeksjonssone, (III) nitrogenfikseringssone, (IV) senescenssone. Symbiotiske celler i sone II inneholder differensierende endosymbionter, mens i sone III er vertscytoplasma fullpakket med lange nitrogenfikserende bakterier. Endosymbionter farget Med Syto9 har grønn fluorescens.
Vekst Av Symbiotiske Celler Involverer Forsterkning Av Vertsgenomet ved Endoredupliseringssykluser
Ekstrem plantecelleforstørrelse kan observeres i både determinate og ubestemte knuter. Cytoplasma av en nitrogen-fikserende symbiotisk celle er vert for ca 50.000 bakterier. For å imøtekomme et så høyt antall endosymbionter vokser vertscellene. I m. truncatula nodules er volumet av nitrogenfikseringscellene 80 ganger større enn for de diploide meristematiske cellene. Veksten av infiserte celler skjer trinnvis i sone II OG er konsekvensen av gjentatt endoreduplikasjon (ER) av genomet uten mitose. I sone II er cellesyklusmaskineriet fortsatt aktivt, men mangelen på mitotiske sykliner hemmer mitose og forvandler de mitotiske syklusene til endoreduplikasjonssykluser (Cebolla et al., 1999). Dette oppnås ved cellesyklusbryteren CCS52A protein som ved ødeleggelsen av de mitotiske sykliner induserer gjentatte runder av genom duplisering som fører til dannelsen av gradvis voksende polyploide celler (Roudier et al.( 2003; Kondorosi og Kondorosi, 2004). I Medicago-arter kan ploidinivåene nå 64C som representerer 64 ganger høyere DNA-innhold sammenlignet med haploide celler (C tilsvarer haploid DNA-innholdet; Vinardell et al., 2003). Ned-regulering AV CCS52A I m. truncatula hadde ingen effekt på primordium dannelse, men var skadelig for nodule differensiering indikerer AT er sykluser og dannelse av store svært polyploide celler er avgjørende for nodule funksjon (Vinardell et al., 2003). Interessant nok er kortikale celler som inneholder am-sopp også polyploide, så vel som nematodeforende gigantiske rotceller (Favery et al., 2002; Genre et al., 2008). På samme måte er insektsymbiotiske celler, bakteriocyttene som har intracellulære endosymbionter, også store og polyploide (Nakabachi et al., 2010). I angiosperm-planter er polyploidi hyppig, og det spesifikke arvelige mønsteret av polyploidi i forskjellige organer, vev og celletyper tyder på at det kan være en viktig kilde til vertscellens spesialiserte fysiologi (Nagl, 1976; Edgar et al ., 2014). Foruten cellevekst, kan flere genkopier, mangel på kromosomkondensasjon bidra til høyere transkripsjonelle og metabolske aktiviteter. Imidlertid antyder forening av polyploidi med forskjellige cellefunksjoner en innvirkning av polyploidi også på arkitekturen av nukleosomer og på epigenomet som styrer aktivering eller undertrykkelse av bestemte genomiske regioner. Følgelig synes polyploid genominnholdet i symbiotiske celler å være en forutsetning for noduledifferensiering og for ekspresjon av de fleste symbiotiske vertsgener (Maunoury et al., 2010).
Forskjellige Skjebner Av Nitrogenfikserende Bakterier
bakteriene frigjort FRA IT er tilstede i vertscytoplasma som organelllignende strukturer, kalt symbiosomer. Bakteriene har ingen direkte kontakt med cytoplasma da de er omgitt av en peribacteroid membran, også kjent som symbiosom membran (sm). Bakteroid, SM og mellomrommet mellom dem utgjør symbiosomet (Catalano Et al., 2004). SM under dannelsen reflekterer sin plasmamembran opprinnelse, senere modifikasjoner av sin sammensetning åpne nye, spesialiserte roller på host-endosymbiont grensesnitt(Limpens et al., 2009; Ivanov et al., 2012; Brear et al., 2013; Sinharoy et al., 2013). Bacteroids multiplisere i voksende vert nodule celler til en viss celle tetthet, tilpasse seg endosymbiotic livsstil og mikroaerobic forhold og modne til nitrogen-fikse bacteroids. Formen og fysiologien til bakterier kan imidlertid være slående forskjellig i de forskjellige belgfrukter. I visse legume verter, nitrogen-feste bacteroids har samme morfologi som dyrkede celler; denne typen bacteroids kan gå tilbake til fritt levende form. I andre foreninger blir bakteriene irreversibelt omdannet til polyploide, forstørrede, ikke-dyrkbare endosymbionter. Disse terminalt differensierte bakteroider kan være langstrakte og til og med forgrenede og 5 – til 10-ganger lengre enn de frie levende cellene eller kan være sfæriske fra 8 til minst 20-ganger forsterket genom avhengig av verten (Mergaert et al ., 2006; Jørgen et al., 2010). Terminal differensiering av bacteroids er vert kontrollert, utviklet seg i flere grener Av Leguminosae familien indikerer vert fordel og sannsynligvis høyere symbiotisk ytelse (Oono et al ., 2010). Terminal bacteroid differensiering er den beste belyst I S. meliloti – m. truncatula symbiosen. I M. Truncatula nodules, de mest synlige hendelsene i terminal bacteroid differensiering forekommer i sone II. Multiplikasjon av bacteroids stopper i midten av sone II hvor celle forlengelse og uniform forsterkning av flere replikene ved endoreduplication sykluser begynne. Langs 2-3 cellelag ved grensen til sone II OG III (kalt interzone) er plutselig vekst av bakterier synlig og når praktisk talt sin endelige størrelse, men nitrogenfiksering finner sted bare i sone III.
Vertspeptider Styrer Bakterioiddifferensiering
Sammenligning av nodule transkriptomer av belgfrukter med reversibel og irreversibel bakterioiddifferensiering avslørte eksistensen av flere hundre små gener som bare var tilstede i genomet til de vertsplantene hvor bakterioiddifferensiering var terminal. I m. truncatula nodule cellene produsere minst 600 nodule-spesifikke symbiotiske peptider (symPEPs). SymPEP-genene aktiveres bare I s. meliloti-infiserte polyploide symbiotiske celler (Kevei et al., 2002; Mergaert et al., 2003), men visse sett på tidligere, andre i de senere stadier av nodule utvikling. En stor del, mer enn 500 gener kode nodule-spesifikke cystein-rik (NCR) peptider (Mergaert et al., 2003; Alunni et al., 2007; Nallu et al., 2014). NCR peptider er rettet mot bacteroids og når deres levering til endosymbionts ble blokkert, bacteroid differensiering ble avskaffet viser at peptidene er ansvarlig for terminal differensiering Av s. meliloti bacteroids (Van De Velde et al., 2010). Den høye sekvensvariasjonen og DE karakteristiske uttrykksmønstrene TIL NCR-gener tyder på mangfold i deres funksjoner, virkningsmåter og bakterielle mål på forskjellige stadier av bakterioid modning (Figur 2). Men hvorfor produserer vertscellen et arsenal Av NCRs? Hva kan være fordelen med et så variert peptidrepertoar? Er det nødvendig for samspill av verten med ulike bakterier? De symbiotiske partnerne Til m. truncatula er S. meliloti og S. medicae, men i jorda er det utallige stammevarianter av begge arter. M. truncatula er også representert av mange forskjellige økotyper og tiltredelser som varierer i antall, sekvenser og uttrykksprofil AV NCR-gener og i deres symbiotiske interaksjoner med forskjellige S. meliloti og S. medicae-stammer (Nallu et al., 2014; Roux et al., 2014). Mens en knute inneholder en enkelt bakterietype, kan de forskjellige nodulene på samme rotsystem ha forskjellige bakteriepopulasjoner. Det er mulig at planten som gjenkjenner de forskjellige endosymbiontene manipulerer dem med et stamme-spesifikt repertoar av peptider. Disse forskjellene kan legge til et ekstra kontrollnivå for verts-symbiont-spesifisitet og dermed for nodulasjonseffektivitet.

FIGUR 2. Differensial ekspresjon av symPEP-gener I m. truncatula noduler. Svart signal: in situ hybridisering, blå signal: gus aktivitet av symPEP promoter-GUS fusjoner i transgene knuter.selv om symPEPs representerer unike peptidklasser, ligner deres strukturer antimikrobielle peptider (Ampere). Forsterkere med bredt spekter av mikrobiell celledrepende aktivitet er oftest kationisk provoserende celledød ved poredannelse, membranforstyrrelser og påfølgende lysis av mikrobielle celler. Det faktum at celledeling evne er definitivt tapt under endosymbiont differensiering indikerer at minst visse symPEPs har antimikrobielle aktiviteter. Behandling av bakterier med syntetiske kationiske NCRs forårsaket faktisk rask og effektiv doseavhengig eliminering av Ulike Gram-negative og Gram-positive bakterier, inkludert viktige humane og plantepatogener (Van De Velde et al., 2010; Tiricz et al., 2013). Denne ex-planta drap effekt korrelert med permeabilization av mikrobielle membraner, men symPEPs i sitt naturlige miljø-i nodule celler-ikke permeabilize bakterielle membraner og ikke drepe endosymbionts. Sannsynligvis er peptidkonsentrasjonene i nodulene betydelig lavere enn de som ble brukt i in vitro-analysene. Videre produseres kationiske peptider sammen med anioniske og nøytrale peptider i samme celle, og mulig kombinasjon av noen få titalls eller hundrevis av peptider med forskjellig ladning og hydrofobicitet kan nøytralisere den direkte bakteriedrepende effekten av kationiske peptider.involvering Av Forsterkere eller AMP-lignende peptider er ikke unikt For Rhizobium-legume symbiose. I weevil Sitophilus produserer de symbiotiske cellene det antimikrobielle peptidet coleoptericin – A (ColA)som provoserer utviklingen av gigantiske filamentøse endosymbionter ved å hemme celledeling og beskytter de nærliggende insektvevene fra bakteriell invasjon (Login et al ., 2011). I dette systemet er et enkelt peptid tilstrekkelig for differensiering av den obligate vertikalt overførte endosymbionten i motsetning til knuter som opererer med hundrevis av symPEPs og kan være vert for utallige belastningsvarianter som deres endosymbionter. I aphid-Buchnera-symbiosen produserer vertscellene også bakteriocyttspesifikke peptider, inkludert cysteinrike peptider (Bcr) som ligner Medicago NCR-peptidene, men funksjonene til disse symbiotiske peptidene er ikke rapportert ennå (Shigenobu og Stern, 2013).
NCR247: Et Eksempel på Multi-Target Host Effector
Transkriptomanalyse Av M. truncatula knuter på ulike stadier av deres utvikling, laser microdissection av nodule regioner, in situ hybridisering, immunolocalization av utvalgte peptider, og symPEP promoter-reporter genet fusjoner i transgene knuter tillate kartlegging virkningen av individuelle peptider i symbiotiske celler fra tidlig infeksjon til sent nitrogen fiksering tilstand. NCR247 uttrykkes i de eldre cellelagene i sone II og i intersonen hvor bakteriell celledeling stopper og bemerkelsesverdig forlengelse av endosymbiontene oppstår (Farkas et al., 2014). Dette lille kationiske peptidet drepte effektivt forskjellige mikrober in vitro, og in silico-analysen indikerte sin ekstreme proteinbindingskapasitet. FITC-merket NCR247 kom inn i bakteriell cytosol hvor dets interaksjoner med mange bakterielle proteiner var mulige. Binding partnere ble identifisert ved behandling Av s. meliloti bakterier eller bacteroids Med StrepII / FLAGG-merket peptider etterfulgt av affinitet kromatografi og identifisering av samspill partnere MED LC-MS / MS Og Western analyse (Farkas et al ., 2014).En av interaktorene var FtsZ celledeling protein som spilte en avgjørende primær rolle i celledeling. En rekke antibiotiske peptider er kjent for å utøve bakteriedrepende eller bakteriostatisk effekt gjennom samspillet Med FtsZ, og hemmer polymerisasjonen og hindrer dermed riktig Z-ring og septumdannelse (Handler et al., 2008). NCR247 ble ko-renset med FtsZ fra bakteriell cytoplasma og ble vist å forstyrre septumdannelse. NCR035 utviser også bakteriedrepende effekt in vitro og produsert i de samme symbiotiske cellene SOM NCR247 akkumuleres ved divisjonsseptumet som indikerer samtidig eller påfølgende virkning av disse peptidene og utvikling av flere vertsstrategier for å hemme endosymbiontproliferasjon. En annen studie viste at ekspresjon av viktige celledeling gener, inkludert gener som kreves For Z-ring funksjon, ble sterkt svekket i celler behandlet AV NCR247 (Penterman et al., 2014). Forbehandling av bakterier med sub-dødelige NCR247 konsentrasjoner avskaffet lokalisering AV FITC-NCR035 til septum og provosert celleforlengelse (Farkas et al., 2014).
Ribosomale proteiner var DE MEST tallrike NCR247 interaksjonspartnerne. NCR247 ble observert å sterkt hemme bakteriell proteinsyntese på en doseavhengig måte både in vivo og in vitro (Farkas et al., 2014). Disse resultatene antydet at EN modus FOR NCR247 peptidvirkningen er bindende for ribosomene både i bakterielle celler og bakterier. Interessant nok ble et endret mønster og redusert kompleksitet av de interagerende proteiner observert i bakteriene. Følgelig var det generelle ekspresjonsnivået for ribosomale proteiner i gjennomsnitt 20 ganger lavere i bakteriene enn i de frie levende cellene med forskjellig relativ overflod av transkripsjoner av individuelle ribosomale proteiner. Ribosomdiversifisering i bakterier kan ha en betydelig rolle ved å bidra til den avanserte oversettelsen av spesifikke proteiner og dermed støtte den spesialiserte, energikrevende fysiologien til svært rikelig nitrogenfikseringsfunksjon.Den GroEL chaperon var også en direkte samspill partner AV NCR247 (Farkas et al., 2014). Ut av 5 GroEL proteiner, GroEL1 Eller GroEL2 er tilstrekkelig for å overleve Mens GroEL1 uttrykt på høyt nivå i nodule er avgjørende for symbiose (Bittner et al ., 2007). Det er nødvendig for full aktivering av nodulasjonsgener og montering av nitrogenasekomplekset. GroEL har ekstrem funksjonell allsidighet ved å samhandle med hundrevis av proteiner. NCR247-GroEL1-interaksjonen kan ha konsekvenser direkte på GroEL, men indirekte også På Groelsubstrater og tilhørende biologiske prosesser. Fravær Av GroEL1 alvorlig påvirket bakteriell infeksjon og vedlikehold og differensiering av bakterier som viser et generelt behov For GroEL1 i alle stadier av nitrogenfikseringsnudelutvikling.involvering Av GroEL og vert peptider i mikrobe-vert interaksjoner er ikke unik For Rhizobium-legume symbiose. I weevil symbiotiske celler koleoptericin-A (ColA) samhandler også Med GroEL (Login et al., 2011). GroEL spiller også en viktig rolle i vedlikehold av endosymbionter (Moran, 1996; Kupper et al., 2014). Som de fleste symbiotiske systemer er ennå uutforsket og høy gjennomstrømning genomisk og proteomisk verktøy er bare nylig tilgjengelig, kan vi bare forutsi at verts peptider-mediert endosymbiont differensiering, likeledes genom forsterkning av vertsceller og terminalt differensierte endosymbionter er generelle strategier for symbiose.
Konklusjon
Symbiotiske og patogene bakterier bruker lignende tilnærminger til å samhandle med sine verter og å overleve i vertsceller, selv om resultatene av disse interaksjonene er påfallende forskjellige. Planter og dyr kan generere medfødte immunresponser mot mikroorganismer ved oppfatningen Av MAMPs (mikroorganisme-associated molecular patterns). Denne oppfatningen resulterer i aktivering av signaleringskaskader og produksjon av antimikrobielle effektorer. AMP – lignende vert peptider som M. truncatula NCR peptider eller weevil ColA antimikrobielle peptid spille sentral og mangesidig roller i å kontrollere multiplikasjon og differensiering av endosymbionts, og dermed begrense tilstedeværelsen av bakterier til symbiotiske celler. Dermed bruker vertsorganismer disse effektorpeptidene til å temme og til og med ansette utvalgte mikrobielle inntrengere for service.
Interessekonflikt
forfatterne erklærer at forskningen ble utført i fravær av kommersielle eller økonomiske forhold som kan tolkes som en potensiell interessekonflikt.
Anerkjennelser
Arbeid i våre laboratorier støttes av Det Europeiske Forskningsrådets» SYM-BIOTICS » Avanserte Tilskudd til É Kondorosi (stipendnummer 269067) og AV TÁ-4.2.2.A-11/1 / KONV-2012-0035 støttet Av Den Europeiske Union og medfinansiert av Det Europeiske Sosialfond.
Abdel-Lateif, K., Bogusz, D., Og Hocher, V. (2012). Rollen av flavonoider i etableringen av planterøtter endosymbioser med arbuscular mycorrhiza sopp, rhizobia og Frankia bakterier. Plantesignal. Oppfør deg. 7, 636–641. doi: 10.4161 / psb.20039
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Alunni, B., Kevei, Z., Redondo-Nieto, M., Kondorosi, A., Mergaert, P., and Kondorosi, E. (2007). Genomic organization and evolutionary insights on GRP and NCR genes, two large nodule-specific gene families in Medicago truncatula. Mol. Plant Microbe Interact. 20, 1138–1148. doi: 10.1094/MPMI-20-9-1138
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Barnett, M. J., Fisher, R. F., Jones, T., Komp, C., Abola, A. P., Jørgensen, F., et al. (2001). Nukleotidsekvens og forventede funksjoner av hele sinorhizobium meliloti pSymA megaplasmid. Proc. Natl. Acad. Sci. Usa 98, 9883-9888. doi: 10.1073 / pnas.161294798
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Bittner, A. N., Foltz, A., Og Oke, V. (2007). Bare en av fem groEL gener er nødvendig for levedyktighet og vellykket symbiose I Sinorhizobium meliloti. J. Bakteriol. 189, 1884–1889. doi: 10.1128 / JB.01542-06
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Brear, E. M., Day, D. A., and Smith, P. M. (2013). Iron: an essential micronutrient for the legume-Rhizobium symbiosis. Front. Plant Sci. 4:359. doi: 10.3389/fpls.2013.00359
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Catalano, C. M., Lane, W. S., and Sherrier, D. J. (2004). Biochemical characterization of symbiosome membrane proteins from Medicago truncatula root nodules. Electrophoresis 25, 519–531. doi: 10.1002/elps.200305711
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Capela, D., Barloy-Hubler, F., Gouzy, J., Bothe, G., Ampe, F., Batut, J.,Et al. (2001). Analyse av kromosomsekvensen av legume symbiont Sinorhizobium meliloti stamme 1021. Proc. Natl. Acad. Sci. Usa 98, 9877-9882. doi: 10.1073 / pnas.161294398
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Cebolla, A., Vinardell, J. M., Kyss, E., Olá, B., Roudier, F., Kondorosi, A.,et al. (1999). Den mitotiske inhibitoren ccs52 er nødvendig for endoreduplikasjon og ploidy-avhengig celleforstørrelse i planter. EMBO J. 18, 4476-4484. doi: 10.1093/emboj/18.16.4476
CrossRef Full Text
Chen, W. M., Moulin, L., Bontemps, C., Vandamme, P., Bé, G. og Boivin-Masson, C. (2003). Legume symbiotisk nitrogenfiksering av β-proteobakterier er utbredt i naturen. J. Bakteriol. 185, 7266–7272. doi: 10.1128 / JB.185.24.7266-7272.2003
CrossRef Full Text
Edgar, B. A., Zielke, N. Og Gutierrez, C. (2014). Endocycles: en tilbakevendende evolusjonær innovasjon for post-mitotisk cellevekst. Nat. Pastor Mol. Celle. Biol. 15, 197–210. doi: 10.1038/nrm3756
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Farkas, a., Mar Hryvni, G., Düõ, H., Gyö, Z., Lima, R. M., Medzihradszky, K. F.,et al. (2014). Medicago truncatula symbiotisk peptid NCR247 bidrar til bakterioid differensiering gjennom flere mekanismer. Proc. Natl. Acad. Sci. U. S. A. 111, 5183-5188. doi: 10.1073 / pnas.1404169111
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Favery, B., Complainville, A., Vinardell, J. M., Lecomte, P., Vaubert, D., Mergaert, P.,et al. (2002). The endosymbiosis-induced genes ENOD40 and CCS52a are involved in endoparasitic-nematode interactions in Medicago truncatula. Mol. Plant-Microbe Interact. 15, 1008–1013. doi: 10.1094/MPMI.2002.15.10.1008
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Franssen, H. J., Vijn, I., Yang, W. C., and Bisseling, T. (1992). Developmental aspects of the Rhizobium-legume symbiosis. Plant Mol. Biol. 19, 89–107. doi: 10.1007
CrossRef Full Text
Fraysse, N., Couderc, F., and Poinsot, V. (2003). Surface polysaccharide involvement in establishing the Rhizobium–legume symbiosis. Eur. J. Biochem. 270, 1365–1380. doi: 10.1046/j.1432-1033.2003.03492.x
CrossRef Full Text
Genre, A., Chabaud, M., Faccio, A., Barker, D. G., and Bonfante, P. (2008). Prepenetration apparat montering forut og spår kolonisering mønstre av arbuscular mycorrhizal sopp innenfor roten cortex Av Både Medicago truncatula Og Daucus carota. Plantecelle 20, 1407-1420. doi: 10.1105 / tpc.108.059014
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Handler, A. A., Lim, J. E. og Losick, R. (2008). Peptidinhibitor av cytokinese under sporulering I Bacillus subtilis. Mol. Mikrobiol. 68, 588–599. doi: 10.1111/j.1365-2958. 2008. 06173.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Ivanov, S., Fedorova, E. E., Limpens, E., De Mita, S., Genre, A., Bonfante, P.,et al. (2012). Rhizobium–legume symbiosis shares an exocytotic pathway required for arbuscule formation. Proc. Natl. Acad. Sci. U.S.A. 109, 8316–8321. doi: 10.1073/pnas.1200407109
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Kevei, Z., Vinardell, J. M., Kiss, G. B., Kondorosi, A., and Kondorosi, E. (2002). Glysinrike proteiner kodet av en nodule-spesifikk genfamilie er involvert i ulike stadier av symbiotisk noduleutvikling I Medicago spp. Mol. Plante Mikrobe Samhandle. 15, 922–931. doi: 10.1094 / MPMI.2002.15.9.922
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Kondorosi, E., Og Kondorosi, A. (2004). Endoreduplisering og aktivering av anafasefremmende kompleks under symbiotisk celleutvikling. FEBS Lett. 567, 152–157. doi: 10.1016 / j.febslet.2004.04.075
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Kondorosi, E., Mergaert, P., Og Kereszt, a. (2013). Et paradigme for endosymbiotisk liv: celledifferensiering Av Rhizobiumbakterier provosert av vertsplantefaktorer. Annu. Rev. Mikrobiol. 67, 611–628. doi: 10.1146 / annurev-micro-092412-155630
Kryssref Fulltekst
Kupper, M., Gupta, Sk, Feldhaar, H., Og Brutto, R. (2014). Allsidige roller av chaperonin GroEL i mikroorganisme-insektinteraksjoner. FEMS Mikrobiol. Lett. 353, 1–10. doi: 10.1111 / 1574-6968.12390
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Limpens, E., Ivanov, s., van Esse, W., Voets, G., Fedorova, E., Og Bisseling, T. (2009). Medicago N2-fikse symbiosomer erverve endocytic identitet markør Rab7 men forsinke oppkjøpet av vacuolar identitet. Plantecelle 21, 2811-2828. doi: 10.1105 / tpc.108.064410
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Logg Inn, F. H., Balmand, S., Vallier, A., Vincent-Moné, C., Vigneron, A., Weiss-Gayet, M.,et al. (2011). Antimikrobielle peptider holder insektendosymbionter under kontroll. Vitenskap 334, 362-365. doi: 10.1126 / vitenskap.1209728
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
MacLean, A. M., Finans, T. M., And Sadowsky, M. J. (2007). Genomer av symbiotiske nitrogen-fikserende bakterier av belgfrukter. Plante Physiol. 144, 615–622. doi: 10.1104/s. 107. 101634
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Maunoury, N., Redondo-Nieto, M., Bourcy, M., Van De Velde, W., Alunni, B., Mcgill, P., et al. (2010). Differensiering av symbiotiske celler og endosymbionter er koblet til to transkriptombrytere i Medicago truncatula nodulation. PLoS EN 5: e9519. doi: 10.1371 / tidsskrift.pone.0009519
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Mergaert, P., Nikovics, K., Kelemen, Z., Maunoury, N., Vaubert, D., Kondorosi, A.,Et al. (2003). En roman familie I Medicago truncatula bestående av mer enn 300 nodule-spesifikke gener som koder for små, utskilte polypeptider med konserverte cystein motiver. Plante Physiol. 132, 161–173. doi: 10.1104/pp.102.018192
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Mergaert, P., Uchiumi, T., Alunni, B., Evanno, G., Cheron, A., Catrice, O.,et al. (2006). Eukaryotic control on bacterial cell cycle and differentiation in the Rhizobium-legume symbiosis. Proc. Natl. Acad. Sci. U.S.A. 103, 5230–5235. doi: 10.1073/pnas.0600912103
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Moran, N. A. (1996). Akselerert evolusjon Og Mullers ratchet i endosymbiotiske bakterier. Proc. Natl. Acad. Sci. Usa 93, 2873-2878. doi: 10.1073 / pnas.93.7.2873
CrossRef Full Text
Moran, N. A., McCutcheon, J. P., Og Nakabachi, A. (2008). Genomikk og evolusjon av arvelige bakterielle symbionter. Annu. Pastor Genet. 42, 165–690. doi: 10.1146 / annurev.genet.41.110306.130119
CrossRef Full Text
Nakabachi, A., Koshikawa, S., Miura, T. og Miyagishima, S. (2010). Genomstørrelse Av Pachypsylla venusta (Hemiptera: Psyllidae) og ploidien av bakteriocytten, den symbiotiske vertscellen som har intracellulære mutualistiske bakterier med det minste cellulære genomet. Bull. Entomol. Res. 100, 27-33. doi: 10.1017 / S0007485309006737
Pubmed Abstrakt / Pubmed Full Tekst / CrossRef Full Tekst
Nagl, W. (1976). DNA endoreduplikasjon og polyteny forstått som evolusjonære strategier. Natur 261, 614-615. doi: 10.1038 / 261614a0
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Nallu, S., Silverstein, K. A., Zhou, P., Young, Nd, Og Vandenbosch, Ka (2014). Mønstre av divergens av en stor familie av nodule cysteinrike peptider i tiltredelser Av Medicago truncatula. Plante J. 78, 697-705. doi: 10.1111 / tpj.12506
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Oldroyd, G. E. (2013). Snakk, venn og skriv inn: signaleringssystemer som fremmer gunstige symbiotiske foreninger i planter. Nat. Rev. Mikrobiol. 11, 252–263. doi: 10.1038/nrmicro2990
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Oono, R., Schmitt, I., Sprent, J. I., and Denison, R. F. (2010). Multiple evolutionary origins of legume traits leading to extreme rhizobial differentiation. New Phytol. 187, 508–520. doi: 10.1111/j.1469-8137.2010.03261.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Penterman, J., Abo, R. P., De Nisco, N. J., Arnold, M. F., Longhi, R., Zanda, M.,et al. (2014). Vertsplantepeptider fremkaller en transkripsjonell respons for å kontrollere Sinorhizobium meliloti – cellesyklusen under symbiose. Proc. Natl. Acad. Sci. U. S. A. 111, 3561-3566. doi: 10.1073 / pnas.1400450111
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Pris, D. R., Duncan, Rp, Shigenobu, S., Og Wilson, Ac (2011). Genom ekspansjon og differensial ekspresjon av aminosyre transportører på bladlus / buchnera symbiotisk grensesnitt. Mol. Biol. Evol. 28, 3113–3126. doi: 10.1093/molbev/msr140
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Roudier, F., Fedorova, E., Lebris, M., Lecomte, P., Györgyey, J., Abad, P.,et al. (2003). The Medicago species A2-type cyclin is auxin regulated and involved in meristem formation but dispensable for endoreduplication-associated developmental programs. Plant Physiol. 131, 1091–1103. doi: 10.1104/pp.102.011122
CrossRef Full Text
Roux, B., Rodde, N., Jardinaud, M. F., Timmers, T., Sauviac, L., Cottret, L.,et al. (2014). En integrert analyse av plante-og bakteriegenuttrykk i symbiotiske rotknuter ved hjelp av laseropptaksmikrodisseksjon koblet TIL RNA-sekvensering. Plante J. 77, 817-837. doi: 10.1111 / tpj.12442
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Shigenobu, S., Og Stern, D. L. (2013). Bladlus utviklet nye utskilte proteiner for symbiose med bakteriell endosymbiont. Proc. Biol. Sci. 280, 20121952. doi: 10.1098 / rspb.2012.1952
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Sinharoy, S., Torres-Jerez, I., Bandyopadhyay, K., Kereszt, A., Pislariu, C. I., Nakashima, J.,et al. (2013). Den c2h2 transkripsjon faktor regulator av symbiosome differensiering undertrykker transkripsjon av sekretorisk vei genet VAMP721a og fremmer symbiosome utvikling I Medicago truncatula. Plantecelle 25, 3584-3601. doi: 10.1105 / tpc.113.114017
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Terpolilli, J. J., Hood, G. A., and Poole, P. S. (2012). What determines the efficiency of N(2)-fixing Rhizobium-legume symbioses? Adv. Microb. Physiol. 60, 325–389. doi: 10.1016/B978-0-12-398264-3.00005-X
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Tiricz, H., Szûcs, A., Farkas, A., Pap, B., Lima, R. M., Maróti, G.,et al. (2013). Antimicrobial nodule-specific cysteine-rich peptides induce membrane depolarization associated changes in Sinorhizobium meliloti. Appl. Env. Microbiol. 79, 6737–6746. doi: 10.1128/AEM.01791-13
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Van de Velde, W., Zehirov, G., Szatmari, A., Debreczeny, M., Ishihara, H., Kevei, Z.,et al. (2010). Plant peptides govern terminal differentiation of bacteria in symbiosis. Science 327, 1122–1126. doi: 10.1126/science.1184057
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Vinardell, J. M., Fedorova, E., Cebolla, A., Kevei, Z., Horvath, G., Kelemen, Z.,et al. (2003). Endoreduplikasjon mediert av anafasefremmende kompleksaktivator CCS52A er nødvendig for symbiotisk celledifferensiering I Medicago truncatula noduler. Plantecelle 15, 2093-2105. doi: 10.1105 / tpc.014373
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Walker, S. A. og Downie, J. A. (2000). Oppføring Av Rhizobium leguminosarum bv. viciae inn rot hår krever minimal Nikk faktor spesifisitet, men påfølgende infeksjon trådvekst krever nodO eller nodE. Mol. Plante-Mikrobe Samhandle.13, 54–62. doi: 10.1094 / MPMI.2000.13.7.754
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text