- Innledning
- Materialer og Metoder
- Plantematerialer og Veksttilstand
- PSI-Og PSII-Målinger
- Elektrokromisk Skiftanalyse
- Photoinhibitory Behandlinger
- Statistisk Analyse
- Resultater
- Diskusjon
- Rollen AV Cef Stimulering Ved Lavt Lys
- Endring I P700 Oksidasjon Ratio Ved Svakt Lys Er Ikke Relatert til pmf
- ETRII Styrer P700 Oksidasjonsforhold Ved Lavt Lys
- Konklusjon
- Forfatterbidrag
- Finansiering
- Interessekonflikt
Innledning
selv om lys er drivkraften til fotosyntese, kan høyt lys forårsake betydelig fotoinhibisjon AV fotosystem II (PSII) for blader av skyggeetablerende planter (Kitao et al., 2000; Barth et al., 2001; Krause et al., 2004; Huang et al., 2015b, 2016b, c). Imidlertid gjenoppretter photoinhibited PSII raskt ved lavt lys (He og Chow, 2003; Zhang Og Scheller, 2004, Allakhverdiev et al., 2005), på grunn av den raske omsetningsgraden Av d1 protein (Aro et al., 1993; Zhang Og Scheller, 2004; Allakhverdiev et al., 2005). FREKVENSEN AV psii-reparasjon ble redusert ved hemming av SYNTESEN av ATP enten VIA PSI eller PSII (Allakhverdiev et al., 2005), noe som indikerer at rask reparasjon av fotodamerte PSII-komplekser trenger en stor MENGDE ATP på kort tid. En tidligere studie indikerte at etter chilling-indusert fotoinhibisjon AV PSII ble syklisk elektronstrøm (CEF) signifikant stimulert under utvinning ved lavt lys (Huang et al., 2010). Det ble foreslått at denne stimuleringen AV CEF ved lavt lys forbedret SYNTESEN AV ATP for rask reparasjon AV PSII (Huang et al., 2010). Imidlertid er det behov for mer bevis for å avklare den spesifikke rollen SOM cef-stimulering ved lavt lys etter PSII-fotoinhibisjon.under CEF sykles elektroner fra ENTEN NADPH eller ferredoksin rundt PSI inn i plastokinonbassenget. Denne elektronoverføringen er koblet til protontranslokasjon og genererer en protongradient over thylakoidmembraner (Δ) (Shikanai og Yamamoto, 2017). I tillegg til Å bidra MED ATP-syntese, er En annen funksjon Av Δ en nedregulering av fotosyntetisk elektrontransport ved å surgjøre thylakoid lumen (Shikanai, 2014, 2016). Denne forskriften innebærer to forskjellige mekanismer: den ene er knyttet til termisk energidisponering og sprer overflødig absorbert lysenergi som varme fra PSII-antenner (Takahashi et al., 2009), og den andre er nedregulering Av Cyt b6 / f kompleks aktivitet og styrer frekvensen av elektronoverføring TIL PSI (Suorsa et al., 2012, 2016; Tikkanen og Aro, 2014). I angiospermer inkludert Arabidopsis thaliana opererer to veier FOR PSI syklisk elektrontransport (Shikanai, 2007). Den første cef-banen er pgr5- / PGRL1-avhengig vei følsom for antimycin A (Munekage et al., 2002; Jørgen et al., 2013), og den andre er avhengig av kloroplast NADH dehydrogenase-lignende (NDH) kompleks (Burrows et al., 1998; Shikanai et al., 1998). Bidraget FRA DEN pgr5- / PGRL1-avhengige banen er mer signifikant I C3-planter. Under høyt lys induserer aktiveringen AV pgr5-/PGRL1-avhengig CEF forsuring av thylakoid lumen og fører dermed til høye nivåer Av p700 oksidasjonsforhold (Suorsa et al., 2012, 2016; Kono et al., 2014; Jørgen et al., 2016). Til sammenligning ble p700 oksidasjonsforholdet ved lavt lys lite påvirket av mangelen I PGR5-/PGRL1-avhengig CEF (Munekage et al., 2002, 2004; Kono et al., 2014). Ved moderat psii fotoinhibition ble stimuleringen AV CEF ved lavt lys ledsaget Av en økning I p700 oksidasjonsrasjonen (Huang et al., 2010). Det er imidlertid uklart om dette økte p700 oksidasjonsforholdet skyldes CEF-stimuleringen.i TILLEGG til CEF spiller elektronoverføring fra PSII (ETRII) en viktig rolle i å påvirke redokstilstanden TIL PSI (Tikkanen et al., 2014; Huang et al., 2016a, c; Suorsa et al., 2016). I pgr5-planter Av A. thaliana vil alvorlig fotoinhibisjon av PSII trolig fungere som den ultimate kontrollen av fotosyntetisk elektronoverføring som tillater vedlikehold Av P700 optimalt oksidert under overflødig lys(Tikkanen et al ., 2014). Nylig, Suorsa et al. (2016) indikerte at pgr5-planter viste høyt p700 oksidasjonsforhold under høyt lys når ETRII ble redusert ved å kombinere knockout-mutasjoner I PsbO1, PsbP2, PsbQ1, PsbQ2 og PsbR loci. I kjølte blader av tobakk, moderat PSII photoinhibition deprimert elektron flyt TIL PSI og deretter økt P700 oksidasjon ratio under ytterligere chilling behandlinger (Huang et al.( 2016a). Videre førte chilling-indusert fotoinhibisjon AV PSII til en depresjon AV ETRII, som ble ledsaget Av en økning I P700-oksidasjonsforholdet ved lavt lys (Huang et al., 2010). Vi hypoteser At p700 oksidasjonsforholdet ved lavt lys primært styres av elektronstrømmen FRA PSII.
i vår forrige rapport, observerte vi at skygge-etablere anlegget Panax notoginseng viste selektiv photoinhibition AV PSII under høy-lys stress. Etter kortvarig eksponering for høylysspenning, ble ETRII ved lavt lys signifikant redusert, MEN CEF ble signifikant stimulert. Samtidig økte P700 oksidasjonsforholdet i stor grad. Våre spesifikke mål var å (1) undersøke om CEF-stimulering ved lavt lys hovedsakelig letter SYNTESEN AV ATP; og (2) avgjøre om økningen I p700 oksidasjonsforhold ved moderat psii fotoinhibisjon er mer relatert til ETRII i stedet FOR CEF-stimuleringen eller endringen i pmf. For å besvare disse spørsmålene ble intakte Blader Av Panax notoginseng behandlet ved 2258 µ fotoner m-2 s-1 i 30 min. Før og etter behandling med høyt lys ble energidistribusjonen I PSII, redokstilstanden TIL PSI og protonmotivkraft (pmf) ved svakt lys på 54 µmol fotoner m-2 s-1 bestemt.
Materialer og Metoder
Plantematerialer og Veksttilstand
i denne studien ble 2 år gamle planter av en skyggeetablerende plante Panax notoginseng (Burkill) Fh Chen ex C. Chow og Wg Huang brukt til eksperimenter. Disse plantene ble dyrket i lysforhold på 10% sollys (med maksimal lysintensitet midt på dagen på omtrent 200 µ fotoner m-2 s-1). Ingen vann-og ernæringsspenninger ble opplevd for disse plantene. 9 uker gamle fullt utvidede blader ble brukt til fotosyntetiske målinger.
PSI-Og PSII-Målinger
Fotosystem i-og Psii-parametere ble målt til 25°c ved samtidig registrering ved Bruk Av Et Dobbelt pam-100-målesystem (Heinz Walz GmbH, Effeltrich, Tyskland). Klorofyllfluorescensparametrene ble beregnet som følger: Fv / Fm = (Fm-Fo) / Fm, Y (II) = (Fm ‘- Fs) / Fm’ (Genty et al., 1989), NPQ = (Fm – Fm’)/Fm’. Fo Og Fm er minimum og maksimum fluorescens etter mørk akklimatisering, henholdsvis. Fs er lys-tilpasset steady-state fluorescens. Fo og Fm ble bestemt etter mørk akklimatisering i 30 min før og etter høylysbehandling. PSI-fotosyntetiske parametere ble målt Ved Dual PAM-100 basert På p700-signal (forskjell på intensiteter på 830 og 875 nm pulsmodulert målelys som når fotodetektoren) (Klughammer Og Schreiber, 2008). P700 + – signalene (P) kan variere mellom et minimalt (p700 fullt redusert) og et maksimalt nivå (P700 fullt oksidert). Maksimumsnivået (Pm) ble bestemt ved bruk av en metningspuls (600 ms og 10000 µ fotoner m-2 s-1) etter pre-belysning med langt rødt lys, og Pm ble brukt til å estimere PSI-aktiviteten. Pm ‘ ble bestemt lik Pm, men med aktinisk lys i stedet for langt rødt lys. Kvanteutbyttet TIL PSI ble beregnet Som Y (I) = (Pm’ – P) / Pm. P700 oksidasjonsforholdet i et gitt aktinisk lys ble beregnet SOM Y (ND) = P / Pm. Kvanteutbyttet AV PSI-ikke-fotokjemisk energidissipasjon på grunn av akseptor-sidebegrensning ble beregnet SOM Y (NA) = (Pm – Pm’)/Pm. Steady state-verdiene FOR Y (II), NPQ, Y(I) og Y(ND) ble målt etter lysakklimatisering ved svakt lys i 20 min.
Fotosyntetiske electron strømme gjennom PSI og PSII ble beregnet som: ETRII = Y (II) ×PPFD ×0.84 ×0.5 (Krall og Edwards, 1992), ETRI = Y (I) ×PPFD×0.84 ×0.5 (Yamori et al., 2011), hvor 0,5 antas å være andelen absorbert lys som når PSI eller PSII, og 0,84 antas å være absorbenten (brøkdelen av det innfallende lyset absorbert av blader). Omfanget AV cef-aktivering ble estimert SOM ETRI / ETRII-forhold (Yamori et al., 2011, 2015). Det bør bemerkes at 1: 1 eksitasjon partisjonering MELLOM PSI og PSII kan være ikke ekte, fordi skygge planter har en tendens til å øke PSII / PSI forholdet (Lunde et al., 2003; Tikkanen et al., 2006; Grieco et al., 2012). VIDERE ble PSI-og PSII-parametere i denne studien målt ved bruk av rødt aktinisk lys (635 nm) levert Av Dual-PAM 100 (Walz, Tyskland). Fordi rødt aktinisk lys favoriserer eksitering AV PSII over PSI, kan eksitasjonsenergidistribusjonen fra LHCII TIL PSII og PSI påvirkes av det røde aktiniske lyset (Tikkanen et al., 2017).
Elektrokromisk Skiftanalyse
ECS-signalet ble overvåket som absorbansendringen ved 515 nm ved Hjelp av En Dual-PAM-100 (Walz, Effeltrich, Tyskland) utstyrt Med En P515/535 emitter-detektor modul (Walz). ECS-signalet ble oppnådd etter 20 min belysning ved 54 µ fotoner m-2 s-1, etterpå BLE ECS-forfallet målt ved å slå av aktinisk lys i 30 s. analysen AV ECS dark interval relaxation kinetics (DIRKECS) ble gjort ved metoden Til Sacksteder et al. (2001) Og Takizawa et al. (2008). Total pmf ble estimert ut fra total amplitude av DET raske forfallet AV ECS-signalet under 300 ms mørkt intervall. Den langsomme avslapningen AV ECS-signalet gjorde det mulig å gjenkjenne bidraget av protongradient over thylakoidmembranene (Δ). Tidskonstanten for FØRSTEORDENS ECS-avslapping (tECS) er omvendt proporsjonal med protonledningsevnen (gH+) av thylakoidmembranen gjennom ATP-syntasen (Sacksteder og Kramer, 2000; Cruz et al., 2005). Som et resultat ble gH+ estimert som den inverse av forfallstidskonstanten .
Photoinhibitory Behandlinger
etter mørk akklimatisering i 30 min ble Fv/Fm og Pm målt i intakte blader. Etterpå ble disse intakte bladene lett tilpasset til 59 µ fotoner m-2 s-1 i 20 min og parametere for klorofyllfluorescens, P700-signal og ECS-signal ble registrert. Deretter ble aktinisk lys endret til 2258 µ fotoner m-2 s-1. Etter eksponering for dette høye lyset i 30 min ble aktinisk lys endret til 59 µ fotoner m-2 s-1 umiddelbart, og fotosyntetiske parametere ble registrert etter lysakklimatisering i 20 min. Endelig ble Fv/Fm og Pm målt etter mørk akklimatisering i 30 min.
Statistisk Analyse
resultatene ble vist som middelverdier for fem uavhengige eksperimenter. Uavhengig t-test ble brukt ved α = 0,05 signifikansnivå for å avgjøre om det var signifikante forskjeller mellom ulike behandlinger. Alle statistiske analyser ble utført MED SPSS 16.0.
Resultater
under fotosyntetisk induksjon ved 59 µ fotoner m-2 s-1 økte ETRII gradvis og nådde en steady state ved omtrent 18 min (Figur 1a). TIL sammenligning var ETRI konstant i denne induksjonsfasen (Figur 1a). Etter utbruddet av dette svake lyset var ETRI mye høyere ENN ETRII. Men etter en 20-min fotosyntetisk induksjon var ETRI lavere enn ETRII. Disse resultatene foreslo aktivering AV CEF i den innledende fasen av induksjon, i samsvar med tidligere studier (Joliot og Joliot, 2002, 2005; Makino et al., 2002). Etter denne fotosyntetiske induksjonen ved svakt lys ble bladene opplyst ved et høyt lys på 2258 µ fotoner m-2 s-1 i 30 min. Interessant nok ble ETRII gradvis redusert under høylysbehandlingen (Figur 1b). I mellomtiden var verdien av ETRI høyere enn ETRII (Figur 1b), noe som indikerer aktiveringen AV CEF under høyt lys. Etter denne høylysbehandlingen ble den maksimale fotooksiderbare P700 (Pm) opprettholdt stabil (Figur 2a). Til sammenligning ble maksimal fluorescensintensitet (Fm) redusert med 40% og maksimal kvanteutbytte AV PSII (Fv/Fm) redusert fra 0,80 til 0,65 (Figur 2B, C). Disse resultatene indikerte selektiv fotoinhibisjon AV PSII i blader Av Panax notoginseng under høyt lysspenning.
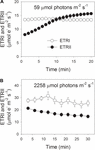
FIGUR 1. Fotosyntetisk elektronstrøm ved lavt lys (A) og høyt lys (B). (A) Mørkakklimatiserte blader ble utsatt for et svakt lys på 59 µ fotoner m-2 s-1 i 20 min. (B) etter belysning ved 59@mol fotoner m-2 s-1 i 20 min for å aktivere elektronvasken i fotosyntese, ble bladene utsatt for et høyt lys på 2258 µ fotoner m-2 s-1 i 30 min. Verdiene er midler ± SE (n = 5).
før behandling med høyt lys ble modne blader opplyst i svakt lys av 59 µ fotoner m-2 s-1 i 20 min for å aktivere fotosyntese, og deretter ble verdier for ETRI, ETRII, Y(ND) og NPQ registrert. Etter behandling med høyt lys i 30 min ble verdier for ETRI, ETRII, Y(ND) og NPQ registrert etter en ny akklimatiseringsfase på 20 min til svakt lys. Før behandling med høyt lys var verdiene FOR ETRI og ETRII ved 59@mol fotoner m-2 s-1 henholdsvis 13,4 og 15,7 µ elektroner m-2 s-1 (Figur 3a, B). Etter behandling med høyt lys var ETRI og etrii ved svakt lys henholdsvis 12,8 og 11,2 µ elektroner m-2 s-1 (Figur 3a, B). Verdien av ETRII ved 59 µ fotoner m-2 s-1 ble redusert med omtrent 30% etter fotoinhibitorisk behandling, noe som indikerer depresjon AV ETRII ved moderat PSII fotoinhibisjon. Før behandling med høyt lys var verdien AV ETRI/ETRII-forholdet ved 59 µ fotoner m-2 s-1 0,85 (Figur 3c). Etter photoinhibitory behandling økte ETRI / ETRII forholdet betydelig til 1,14(Figur 3C). Disse resultatene foreslo stimulering AV CEF ved lavt lys ved PSII photoinhibition (Yamori et al., 2011, 2015).
etter høylysbehandlingen endret ikke steady-state-verdien Av Y(I) ved 59 µ fotoner m-2 s-1 (Figur 4A). Interessant, y (NA) betydelig redusert fra 0,36 til 0,23 (Figur 4B), OG Y(ND) betydelig økt fra 0,1 til 0,25 (Figur 4C). Disse resultatene indikerte endring i redoks tilstand AV PSI ved svakt lys etter fotoinhibitory behandling. I mellomtiden endret verdien AV NPQ litt (Figur 4D). Som et resultat hadde den moderate psii photoinhibition forskjellige effekter På P700 oksidasjonsforhold og NPQ ved lavt lys.
for å avklare om økningen I Y(ND) skyldes en økning i pmf, ble de elektrokromiske skiftsignalene ved 54 µ fotoner m-2 s-1 bestemt etter lysakklimatisering i 20 min før og etter høylysbehandlingen. Interessant nok ble den totale pmf betydelig redusert med 18% etter høylysbehandling (Figur 5a), Men Δ-nivået endret seg ubetydelig (Figur 5b). Fordi dannelsen av pmf kan påvirkes av thylakoidprotonledningsevne, ble protonledningsevnen (gH+) av thylakoidmembranen ved dette svake lyset også undersøkt før og etter høylysbehandling. Resultatet viste at gH+ ikke endret seg med høylysbehandling (Figur 5c), noe som tyder på at høylysbehandlingen knapt påvirket aktiviteten til kloroplast ATP-syntasen ved lavt lys. Ettersom reduksjonen i ETRII på 30% var ledsaget av en reduksjon på 18% i pmf, kompenserte stimuleringen av CEF ved svakt lys delvis for dannelsen av pmf og Δ.

FIGUR 5. Verdier for total protonmotivkraft (pmf) (A), protongradienten (Δ) over thylakoidmembranen (B) Og protonledningsevnen (gH+) av thylakoidmembranen (C) ved svakt lys. Alle parametere ble målt etter tilpasning ved 54 µ fotoner m-2 s-1 i 20 min. Verdiene er midler ± SE (n = 5). Stjerne indikerer en signifikant (P < 0,05) endring etter behandling med høyt lys.
Diskusjon
Rollen AV Cef Stimulering Ved Lavt Lys
DET har blitt indikert AT CEF spiller viktige roller i å opprettholde fotosyntese og plantevekst (Yamori og Shikanai, 2016). Under høyt lys antas CEF nå å være avgjørende for å balansere ATP/NADPH-energibudsjettet, samt for å beskytte PSI og PSII mot skade (Munekage et al., 2002, 2004; Takahashi et al., 2009; Suorsa et al., 2012, 2016; Walker et al., 2014; Huang et al.( 2015a, 2017b). VED lav lysintensitet spiller CEF en viktig rolle i å optimalisere fotosyntetisk CO2-assimilering sannsynligvis via tilførsel av ekstra ATP (Yamori et al., 2011, 2015; Nishikawa et al., 2012; Huang et al.( 2015a). Hovedrollen TIL CEF er modulert fleksibelt som svar på lysintensitet. SPESIELT bidrar CEF hovedsakelig til å balansere ATP / NADPH – energibudsjettet under submettende lysintensiteter, men beskytter hovedsakelig fotosyntetisk apparat mot fotoinhibisjon når det utsettes for metning av lysintensiteter via forsuring av thylakoid lumen (Huang et al.( 2015a). Interessant, Huang et al. (2010) fant AT CEF ble signifikant stimulert ved lavt lys etter chilling-indusert fotoinhibisjon AV PSII og antydet at DENNE CEF-stimuleringen hovedsakelig forbedret SYNTESEN AV ATP for rask reparasjon AV PSII. Imidlertid var det behov for flere bevis for å støtte denne hypotesen. I denne studien observerte vi at verdien AV ETRI / ETRII ved lavt lys økte betydelig (Figur 3C), noe som indikerer stimuleringen AV CEF rundt PSI (Huang et al., 2011, 2012, 2017a, B; Yamori et al., 2011, 2015). Samtidig ble amplituden til pmf signifikant redusert og Δ falt noe. Disse resultatene indikerte at stimuleringen AV CEF ikke utløste Den Δ-avhengige nedreguleringen av fotosyntetisk elektrontransport. Med andre ord lette denne stimuleringen AV CEF ved lavt lys hovedsakelig SYNTESEN AV ATP.Selv om protonledningsevnen til kloroplast-ATP-syntasen ikke ble påvirket av høylysbehandling, resulterte den mindre størrelsen på pmf i en reduksjon i DEN faktiske FREKVENSEN AV ATP-syntese. Satsene FOR PSII-reparasjon og fotosyntetisk CO2-assimilering ved lavt lys er hovedsakelig begrenset AV ATP-syntese (Allakhverdiev et al., 2005; Jørgen et al., 2011, 2015; Nishikawa et al., 2012). FORDI ETRII betydelig redusert PÅ PSII fotoinhibition, frekvensen AV ATP syntese via ETRII ble bemerkelsesverdig redusert. FOR å balansere ATP / NADPH-forholdet som kreves av primær metabolisme og rask reparasjon AV PSII, er det nødvendig med en fleksibel mekanisme for å gi ekstra ATP. UNDER denne tilstanden, ETRII redusert med 30% , men den totale pmf redusert med bare 18%. Disse resultatene indikerer at stimuleringen AV CEF kompenserte for reduksjonen I ETRII-avhengig dannelse av pmf, og økte SYNTESEN AV ATP. Den raske reparasjonen av fotodamert PSII er avhengig AV ATP-syntese (Allakhverdiev et al., 2005). Accelerasjon av CEF rundt PSI økte intracellulær KONSENTRASJON AV ATP, og dermed akselerere frekvensen AV PSII-reparasjon I Synechocystis (Allakhverdiev et al., 2005). Til sammen foreslår vi at stimuleringen AV CEF ved lavt lys spiller en viktig rolle i rask reparasjon AV PSII-aktivitet via en ekstra ATP-syntese.
Endring I P700 Oksidasjon Ratio Ved Svakt Lys Er Ikke Relatert til pmf
et interessant fenomen er At p700 oksidasjon ratio ved svakt lys betydelig økt ved moderat PSII photoinhibition (Huang et al., 2010), som også er vist i denne studien. Under forhold hvor absorbert lys overstiger kravene til fotosyntese, REGULERER CEF-avhengig generasjon Av Δ ned-aktiviteten Til Cyt b6/f kompleks aktivitet og styrer elektronstrømmen FRA PSII TIL PSI (Suorsa et al., 2012, 2016; Shikanai, 2014, 2016), optimaliserer redoks-tilstanden Til P700 i PSI og minimerer ROS-produksjonen i PSI under fotosyntese. I pgr5-planter Av A. thaliana reduseres P700 med elektroner i lyset på grunn av tap av pmf-generasjon. Men i pgr5-mutanten som akkumulerer flavodiironprotein, ble nivået av pmf også gjenopprettet til villtype-nivået, Og Dermed ble P700 oksidert som i villtype (Yamamoto et al., 2016). I klorofyll b-mangelfull hvete mutant linjer, utilstrekkelig thylakoid proton gradient fører til over-reduksjon AV PSI akseptor side og dermed PSI photoinhibition under høy lys eller høy temperatur (Brestic et al ., 2015, 2016). Nylig rapporterte noen studier at den økte aktiviteten til kloroplast ATP-syntasen svekket dannelsen av pmf og forårsaket overreduksjonen av fotosyntetisk elektrontransportkjede, noe som resulterte i fotodamasje AV PSI under høyt lys og svingende lys (Kanazawa et al., 2017; Jørgen et al., 2017). Disse rapportene støtter pmfs kritiske rolle i å opprettholde p700 optimalt oksidert under overflødig lysenergi.
våre nåværende resultater indikerte at etter høylysbehandlingen i 30 min, ble den totale pmf dannet ved dette svake lyset betydelig redusert og dannelsen av Δ Endret noe (Figur 5A,B). I mellomtiden økte p700 oksidasjonsforholdet betydelig fra 0,1 (før behandling) til 0,25 (Etter behandling) (Figur 4C). Disse resultatene tyder sterkt på at denne økningen I P700 oksidasjonsforhold ikke kan forklares med endringen i pmf, som i stor grad er forskjellig fra korrelasjonen MELLOM Y(ND) og pmf under høyt lys (Yamamoto et al., 2016; Ingeborg et al., 2017). Derfor er reguleringseffekten av pmf på p700 oksidasjonsforhold minimal ved lavt lys, men blir spesielt viktig under høyt lys eller svingende lys. Under høyt lys er en høy verdi AV Y(ND) vanligvis ledsaget AV en høy verdi AV NPQ (Munekage et al., 2002, 2004; Kono et al., 2014; Zivcak et al., 2015), på grunn av økningen i pmf. Forringelsen av Δ dannelse fører til reduserte nivåer AV Y (ND) og NPQ under høyt lys (Suorsa et al., 2012; Kono et al., 2014; Jørgen et al., 2016; Kanazawa et al., 2017). Induksjonen AV NPQ ved lavt lys bestemmes hovedsakelig av nivået av lumensurering. Etter behandling med høyt lys endret ikke graden av forsuring av lumen (Figur 5b), OG NPQ forble stabil (Figur 4D). TIL sammenligning økte Y(ND) betydelig. Disse resultatene indikerer at VED lavt lys ble Y(ND) og NPQ kontrollert av ulike reguleringsmekanismer.
ETRII Styrer P700 Oksidasjonsforhold Ved Lavt Lys
PSI blir over-redusert bare når elektronstrømmen fra PSII overskrider kapasiteten TIL PSI elektron akseptorer for å takle elektronene. Når elektronstrømmen TIL PSI er begrenset, ER PSI ekstremt tolerant mot lysspenning i pgr5-plantene Til A. thaliana (Suorsa et al., 2012, 2016; Tikkanen et al., 2014). For skygge-etablering anlegget Psychotria rubra, PSI aktivitet var insusceptible til høy-lys stress i nærvær AV DCMU (Huang et al.( 2016c). I kjølte blader av tobakk tillot moderat PSII-fotoinhibisjon vedlikehold Av P700 optimalt oksidert og deretter beskyttet PSI-aktivitet mot ytterligere fotodamasje (Huang et al.( 2016a). Disse resultatene indikerte DEN viktige rollen ETRII i å kontrollere redoks tilstand AV PSI under høy lys eller chilling-lys påkjenninger. Våre nåværende resultater viste at den moderate psii-fotoinhibisjonen førte til en betydelig depresjon AV ETRII ved lavt lys. I mellomtiden endret PSI-aktiviteten og aktiviteten til kloroplast-ATP-syntasen noe, og den totale pmf og Δ økte ikke. Som et resultat var reduksjonen I ETRII ikke forårsaket av Den Δ-avhengige fotosyntetiske kontrollen via Cyt b6/f-komplekset, men ble sannsynligvis indusert av reduksjonen I PSII-aktivitet (Tikkanen et al., 2014). Den reduserte tilførselen av elektroner fra PSII TIL PSI førte til økt Nivå Av p700 oksidasjon. Således ble p700-oksidasjonsforholdet ved lavt lys i stor grad kontrollert AV ETRII.
Konklusjon
vi fant at den selektive fotoinhiberingen AV PSII induserte en stimulering AV CEF og en økning I P700 oksidasjonsforhold ved lavt lys. Stimuleringen av CEF utløste ikke Den Δ-avhengige nedreguleringen av fotosyntetisk elektrontransport. Som et resultat lette denne stimuleringen AV CEF ved lavt lys hovedsakelig SYNTESEN AV ATP, noe som er viktig for rask reparasjon av fotodamert PSII. Økningen I P700-oksidasjonsforholdet kunne ikke forklares med endringen I Δ-avhengig fotosyntetisk kontroll ved Cyt b6/f-komplekset, men var hovedsakelig forårsaket av redusert tilførsel av elektroner fra PSII til PSI.
Forfatterbidrag
WH, S-BZ og TL utviklet og designet forskningen. WH og Y-JY gjennomførte forsøkene. WH, Y-JY, S-BZ og TL analyserte dataene. WH skrev det første utkastet til manuskriptet, som ble intensivt redigert av alle forfatterne.
Finansiering
denne studien ble støttet Av National Natural Science Foundation Of China (31670343), Youth Innovation Promotion Association Of Chinese Academy Of Sciences (2016347), Og De Store Vitenskap og Teknikk Programmer I Yunnan-Provinsen (2016zf001).
Interessekonflikt
forfatterne erklærer at forskningen ble utført i fravær av kommersielle eller økonomiske forhold som kan tolkes som en potensiell interessekonflikt.
Kono, M., Noguchi, K., Og Terashima, I. (2014). Roller av den sykliske elektronstrømmen rundt PSI (CEF-PSI) og O2-avhengige alternative veier i regulering av fotosyntetisk elektronstrøm i kortsiktig svingende lys I Arabidopsis thaliana. Plantecelle Physiol. 55, 990–1004. doi: 10.1093 / pcp / pcu033
PubMed Abstrakt / CrossRef Full Tekst / Google Scholar
Shikanai, T. (2016). Regulatorisk nettverk av protonmotivkraft: bidrag av syklisk elektrontransport rundt fotosystem I. Photosynth. Res. 129, 253-260. doi: 10.1007 / s11120-016-0227-0
PubMed Abstract | CrossRef Full Text/Google Scholar
Yamori, W., Makino, A., Og Shikanai, T. (2016). En fysiologisk rolle syklisk elektron transport rundt photosystem jeg i å opprettholde fotosyntese under varierende lys i ris. Sci. Rep. 6: 20147. doi: 10.1038/ srep20147
PubMed Abstract | CrossRef Full Text / Google Scholar
Yamori, W., Og Shikanai, T. (2016). Fysiologiske funksjoner av syklisk elektron transport rundt photosystem jeg i å opprettholde fotosyntese og plantevekst. Annu. Rev. Plante Biol. 67, 81–106. doi: 10.1146 / annurev-arplant-043015-112002
PubMed Abstract | CrossRef Full Text/Google Scholar
Yamori, W., Shikanai, T., Og Makino, A. (2015). Photosystem i syklisk elektronstrøm via kloroplast NADH dehydrogenase-lignende kompleks utfører en fysiologisk rolle for fotosyntese ved lavt lys. Sci. Rep. 5:13908. doi: 10.1038 / srep13908
PubMed Abstract / CrossRef Full Text / Google Scholar
Zhang, Sp, Og Scheller, H. V. (2004). Photoinhibition av photosystem i ved chilling temperatur og påfølgende utvinning I Arabidopsis thaliana. Plantecelle Physiol. 45, 1595–1602. doi: 10.1093 / pcp / pch180
PubMed Abstrakt / CrossRef Fulltekst / Google Scholar