- Introdução
- método
- participantes
- A avaliação da criatividade
- Torrance Test of Creative Thinking (Circles Sub-Scale)
- Alternate Uses Task (AUT)
- Pretest
- a Análise Estatística
- Resultados
- o componente de fluência
- o componente flexível
- A componente de originalidade
- diferenças de género
- discussão
- Declaração de conflito de interesses
Introdução
Criativa cognição desempenha um papel importante nas artes, na invenção e a inovação, bem como na vida cotidiana (Runco e Richards, 1997). Embora a criatividade tenha sido considerada uma capacidade humana única, comportamentos espontaneamente criativos (e.g., criando novas ferramentas entre primatas e aves) tem sido mostrado para ocorrer em não-humanos também (Byrne e Bates, 2007), atestando ainda mais as profundas raízes evolutivas e biológicas da criatividade. A criatividade tem sido definida como a capacidade de produzir respostas que são simultaneamente novas (ou seja, originais, raras e inesperadas) e adequadas (ou seja, adaptativas e úteis de acordo com as restrições de Tarefas) (Sternberg e Lubart, 1999). Uma das abordagens psicométricas para medir a criatividade envolve tarefas de pensamento divergente (DT) nas quais os participantes são convidados a responder a um dado problema com múltiplas soluções (Dietrich e Kanso, 2010). Os testes de DT medem geralmente vários aspectos da criatividade, incluindo fluência criativa, flexibilidade e originalidade (Torrance, 1974). Assim, embora não sejam sinónimos de criatividade, as tarefas DT fornecem medidas estruturadas e objetivas de criatividade (Sternberg e Lubart, 1999; Jung et al., 2009). Notavelmente, as pontuações nas tarefas DT têm sido mostradas como positivamente correlacionadas com medidas ecologicamente válidas de realização criativa (Carson et al., 2005), bem como com criatividade auto-avaliada (Furnham e Bachtiar, 2008).estudos anteriores apontaram para o envolvimento do sistema dopaminérgico (DA) na criatividade (Heilman et al., 2003; Flaherty, 2005; Takeuchi et al., 2010). Takeuchi et al. (2010) encontrou diferenças individuais na criatividade, como medida por DT, a ser positivamente correlacionada com a matéria cinzenta nas regiões do sistema, incluindo o córtex pré-frontal dorsolateral, gânglios basais bilaterais, substantia nigra, e a área ventral tegmental. Além disso, vários estudos genéticos demonstraram uma relação entre a TD e a neurotransmissão dopamina (Reuter et al., 2006; de Manzano et al., 2010; Runco et al., 2011). Por exemplo, Reuter et al. (2006) descobriu que a criatividade, medida por tarefas DT envolvendo tanto a criatividade figural quanto a verbal, estava significativamente associada a polimorfismos do gene receptor dopamina D2 (DRD2). Além disso, vários estudos descobriram que a dopamina cortical está envolvida na flexibilidade cognitiva (Frank, 2005; Cools, 2008; Garcia-Garcia et al., 2010), um dos principais componentes da DT.um gene candidato à criatividade é o gene receptor dopamina D4 (DRD4). O receptor DRD4 é um dos cinco receptores dopaminérgicos e desempenha um papel importante na mediação da sinalização sináptica dopamina. O gene é caracterizado por um número variável de 48 pares de bases de repetições tandem (48-bp VNTR) localizado na região de codificação do terceiro exon. DRD4 48-bp polimorfismo VNTR varia de 2 a 11 repetições entre indivíduos (Asghari et al., 1995; Cravchik e Goldman, 2000), onde a 4-repetição (4R) é a repetição mais comum em populações caucasianas e a 7-repetição (7R) é a segunda variante mais comum (Chang et al., 1996; Ding et al., 2002). Curiosamente, o 7R tem sido anteriormente associado com comportamentos criativos da vida real, como o traço de personalidade que busca novidades (Ebstein et al., 1996). Na verdade, a novidade-a busca—a tendência para a atividade exploratória—é considerada uma das características das pessoas criativas (Chavez-Eakle et al., 2006; Drago et al., 2009). Embora a evidência de uma associação entre DRD4 7R e novelty-seeking seja inconsistente, como evidenciado por vários relatórios de meta-análise (Kluger et al., 2002; Munafò et al., 2008), muitos estudos encontraram uma associação significativa entre a busca pela novidade e o alelo 7R do DRD4 (Ebstein et al., 1996; Benjamin et al., 2000; Keltikangas-Järvinen et al., 2002; Becker et al., 2005). Roussos et al. (2009) have recently suggested that differences in measuring novelty-seeking using self-report scales may account for the inconsistent results between 7R polymorphism and novelty-seeking. Em linha com isso, tem sido repetidamente encontrado que o sistema DA desempenha um papel importante no traço de personalidade da busca de novidade (Flaherty, 2005; Schweizer, 2006), atestando ainda o papel potencial do DRD4 na criatividade.
no entanto, em contraste com o papel do DRD4 48-bp VNTR 7R na busca de novidades, evidências recentes indicam que 7R está realmente associado com a flexibilidade cognitiva diminuída, um dos principais aspectos da criatividade (Strobel et al., 2004; Congdon et al., 2008). Flexibilidade envolve a capacidade de fazer uma resposta alternativa após inibir com sucesso uma resposta atual. Até à data, os resultados relativos à associação entre inibição e DRD4 são mistos. Congdon et al. (2008) found that participants with the 7R allele of the DRD4 exhibit higher stop-signal reaction time (SSRT) on a go/no-go task, reflecting poor inibitory control, while Forbes et al. (2009) failed to find such an effect on the Barratt impulsive Scale (BIS), a self-report to measure impulsivity. Por outro lado, Colzato et al. (2010) demonstrou que esses efeitos mistos podem ser devidos ao fato de que estudos anteriores abordaram a impulsividade como um processo monolítico, enquanto a impulsividade pode realmente ser dividida em tipos funcionais e disfuncionais de acordo com o inventário de impulsividade de Dickman (DII; Dickman, 1990). Impulsividade disfuncional é a tendência a agir sem previsão nos casos em que tal ação é inadequada, enquanto a impulsividade funcional é uma tendência semelhante implementada em situações apropriadas (Colzato et al., 2010). Colzato et al. (2010) descobriu que indivíduos com DRD4 7R, que está associado a níveis mais elevados de ESTRIATAL DA, exibiam maior impulsividade disfuncional. Vários estudos meta-analíticos apontaram para o alelo 7R do DRD4 como um alelo de risco para a perturbação da hiperactividade défice de atenção (ADHD; Faraone et al., 2001; DiMaio et al., 2003), um transtorno caracterizado por alta impulsividade disfuncional (Young et al., 2007). Além disso, a relação entre ADHD e criatividade foi investigada em vários estudos (Shaw e Brown, 1990).; Healey and Rucklidge, 2006), though so far these studies have yellowed unclear results. Enquanto alguns acharam a TDAH e sua sintomatologia positivamente correlacionada com a criatividade (Healey e Rucklidge, 2006; White e Shah, 2011), outros relataram uma tendência oposta (Funk et al., 1993; Healey and Rucklidge, 2005, 2008). Curiosamente, a TDAH está comportamentalmente associada a incapacidades no controle executivo, incluindo mudança de conjunto de atenção (Boonstra et al., 2005), and flexibility (Barkley et al., 1997; Sargento et al., 2002), ambos importantes para a flexibilidade cognitiva, uma dimensão central da criatividade (Dietrich, 2004; Durstewitz e Seamans, 2008). Estes resultados indicam que os indivíduos com o alelo 7R do DRD4 podem apresentar níveis mais baixos de criatividade e menor flexibilidade nas tarefas DT em particular. Assim, o DRD4 exon III VNTR é um candidato biologicamente plausível para contribuir para as diferenças individuais na criatividade.colectivamente, parece que, por um lado, o alelo 7R, como alelo de risco para funções executivas comprometidas, flexibilidade e mudança de Conjuntos, pode estar associado a pouca criatividade. Por outro lado, a associação entre o alelo 7R e a maior busca de novidades indica que os indivíduos com o alelo 7R podem realmente exibir maior flexibilidade e criatividade.
O estudo actual foi concebido para explorar o papel complexo do DRD4 7R na formação parcial da criatividade humana. Para melhor caracterizar esta relação, avaliamos dois tipos de testes DT (figural e verbal). A pontuação de cada tarefa foi dividida de acordo com as três principais dimensões de DT, nomeadamente criativo, a fluência, flexibilidade e originalidade, e cada dimensão foi analisada separadamente para testar a hipótese de que DRD4 diferencialmente afeta cada dimensão de criatividade.
método
participantes
a amostra compreendia 185 estudantes da Universidade de Haifa em Israel (112 mulheres). Todos os participantes foram caucasianos (auto-notificados), com uma idade média de 24, 5 (DP ± 2, 1). Os participantes foram recrutados através de anúncios publicados na Universidade. Todos os participantes receberam voluntários.
A avaliação da criatividade
a criatividade foi avaliada utilizando dois testes DT, uma sub-escala do sub-teste figural do teste Torrance de pensamento criativo (Torrance, 1974), e o Alternate Uses Task (AUT; Guilford et al., 1978). Ambos os testes incluíram as três dimensões principais da DT, nomeadamente flexibilidade, originalidade e fluência.
Torrance Test of Creative Thinking (Circles Sub-Scale)
Participants were given a page on which 30 identical circles were drawn. Eles foram convidados a desenhar o maior número possível de objetos significativos dentro de um prazo de 10 minutos, onde cada desenho deve incluir pelo menos um círculo. A pontuação incluía fluência( número de respostas produzidas), flexibilidade (número de categorias) e originalidade, calculada de acordo com a pontuação das respostas originais, conforme detalhado nos testes de Torrance do Creative Thinking scoring guide (Torrance, 1974).
Alternate Uses Task (AUT)
participantes receberam uma lista de cinco objetos comuns e pediram para listar o maior número possível de usos alternativos para cada objeto, dentro de um prazo de 10 minutos. O uso diário mais comum foi indicado entre parênteses. Os objetos eram: sapato( uso comum: desgaste no pé); Botão( uso comum: fechamento de coisas); lápis (uso comum: desenho ou escrita); pneu (uso comum: roda de carro); e copo de bebida (uso comum: contém líquido). Só foram contabilizadas e incluídas as respostas que não reiteraram as utilizações comuns indicadas. Tal como no teste de Torrance, a pontuação incluía fluência, flexibilidade e originalidade. Uma vez que não há diretrizes para a pontuação das respostas originais na UAE, as respostas originais foram definidas como respostas estatisticamente infrequentes de acordo com um pré-teste realizado em nosso laboratório.
Pretest
com a finalidade de criar um critério Válido de frequência de Resposta, Um grupo de 100 participantes saudáveis que não participaram neste estudo completaram a CTA. Para cada objeto, uma lista de todos os usos possíveis foi coletada de todos os participantes. Uma medida de infrequência estatística foi calculada com base nesta lista, a fim de avaliar a pontuação de originalidade para cada resposta e, posteriormente, para cada participante. As respostas foram atribuída uma pontuação de zero se 5% ou mais dos participantes listados a um determinado uso, a pontuação de um se entre 2% e 4,99% das participantes listados, e uma pontuação de dois, se menos de 1.99% listados a utilizar. Uma pontuação de originalidade média foi calculada para cada participante de acordo com estas pontuações estatísticas de infrequência.a extracção e genotipagem do ADN de 20 ml de amostras de Elixir bucal foram extraídas utilizando o “Master Pure kit” (epicentro, Madison, WI). O DRD4 48-bp VNTR foi caracterizado por um procedimento de amplificação PCR com os seguintes iniciadores: F5′ – CTT CCT ACC CTG CCC GCT CAT GCTGCT CTA CTG – 3′ e R5′ – ACC ACCACC GGC AGG ACC CTC ATG GCC TTG CGC TC – 3. As reacções PCR foram conduzidas utilizando 5 µl Master Mix( termo científico), 2 µl primers (0,5 µM), 0,6 µl Mg/Cl2 (2,5 mM), 0.4 µl DMSO 5% e 1 µl de água para um total de 9 µl volume total, e um adicional de 1 µl de ADN genómico foi adicionado à mistura. Todas as reacções PCR foram efectuadas num termociclador Biometra T1 (Biometra, Güttingem, Alemanha). A reacção PCR foi a seguinte: fase de pré-aquecimento a 94,0 ° C durante 5 minutos, 34 ciclos de desnaturação a 94,0°C durante 30 segundos, reanálise a 55°C durante 30 segundos e extensão a 72°C durante 90 segundos. a reacção manteve-se a 72°C durante 5 minutos. A mistura de reacção foi então electroforizada num gel de agarose a 3% (AMRESCO) com brometo de etídio para detectar genótipos.
a distribuição da frequência do genótipo foi de acordo com o equilíbrio de Hardy-Weinberg (qui-quadrado = 0, 01, p valor = 0, 92).
a Análise Estatística
Genótipo foi classificado de acordo com a presença ou ausência do alelo 7R do DRD4 (7 vs. no7), como em linha com os relatórios anteriores na literatura (Ding et al., 2002). As diferenças de grupo nas pontuações da criatividade foram analisadas estatisticamente usando ANOVAs, com a presença (7) ou ausência (no7) do alelo DRD4 7R como variável independente. Para examinar os diferentes aspectos da DT (fluência, flexibilidade, originalidade), realizamos uma análise multivariada da variância (MANOVA) separadamente para cada componente.
Resultados
Para confirmar que os dois grupos (7, no7) não foram significativamente diferentes em termos de idade e de educação, realizamos independente de testes t para comparar os dois grupos (ver Tabela 1 médias e desvios-padrão). Esta análise revelou que os dois grupos não diferiam em termos de idade ou anos de educação. Testes não paramétricos (Mann–Whitney) revelaram diferenças significativas na distribuição de frequência para o sexo (Z = 2.11, p < 0,05).
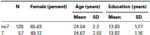
a Tabela 1. Dados relativos à idade, sexo e educação para o genótipo DRD4 7R, SD: desvio-padrão.
as pontuações médias das medidas de criatividade utilizadas nas duas tarefas estão resumidas na Tabela 2.

Tabela 2. Meios e erro-padrão (SE) das duas tarefas de criatividade utilizadas: AUT e Torrance.
o componente de fluência
uma ANOVA multivariada indicou um efeito geral do genótipo DRD4 7R para o componente de fluência, F (2,182) = 3,25, p < 0,05. Dada a importância do teste global, foram examinados os principais efeitos não variáveis. Foram obtidos efeitos principais univariados significativos para o genótipo DRD4 para a fluência AUT F (1,183) = 4, 95, p < 0.05 (Alelo 7R exibindo menor fluência em comparação com o grupo no7, figura 1A), mas não atingiu o nível de significância para a fluência Torrance F(1,183) = 2,95, p = 0,087.
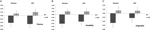
a Figura 1. A criatividade pontua nas tarefas da AUT e da Torrance separadas pela fluência (a), flexibilidade (B) e originalidade (C). Os meios foram transformados em Z partituras.
o componente flexível
a ANOVA multivariada das medidas de flexibilidade exibiu um efeito geral para o genótipo F DRD4 7R (2,182) = 4.38, p < 0,05. Significativo univariada efeitos principais para DRD4 genótipo foram obtidos para AUT flexibilidade F(1,183) = 4.57, p < 0,05 e Torrance flexibilidade F(1,183) = 6.95, p < 0.01. Como mostrado na figura 1B, em ambos os casos os indivíduos do grupo Alelo 7R exibiram menor flexibilidade em comparação com os indivíduos do grupo no7 (Quadro 2).
A componente de originalidade
a ANOVA multivariada das medidas de originalidade não apresentou quaisquer efeitos significativos, F (2,182) = 2, 06, p = 0.13, Embora as pontuações indicassem menor originalidade para o alelo 7R do DRD4 (ver Figura 1C, Quadro 2 para mais pormenores).
diferenças de género
discussão
DT tarefas exigem que os participantes forneçam múltiplas soluções para um dado problema. Nas tarefas aqui utilizadas, os participantes foram obrigados a pensar em muitos usos alternativos possíveis para itens cotidianos (na AUT), e a desenhar vários desenhos incorporando uma forma específica (círculo) (na tarefa Torrance). Os resultados aqui apresentados demonstram que os indivíduos com o alelo DRD4 7R exibem menor criatividade como medida por DT. Considerando que o alelo 7R tem sido associado a impulsividade disfuncional e fraca inibição (Congdon et al., 2008; Dreber et al., 2009), o fato de que indivíduos com o alelo 7R exibem menor criatividade pode ser explicado por sua incapacidade de suprimir ou inibir respostas óbvias (comuns), como indicado por escores de originalidade mais baixos. Uma menor flexibilidade e pontuações de fluência podem indicar que uma vez dada uma resposta comum, torna-se mais difícil mudar para uma nova categoria (como indicado pelas pontuações de baixa flexibilidade), levando a menos respostas (como indicado pela pontuação de baixa fluência). Outras evidências para a relação entre criatividade e impulsividade podem ser vistas na associação do alelo 7R com TDAH (Li et al., 2006), bem como com o aumento da impulsividade em pacientes com TDAH (Langley et al., 2004). Curiosamente, a criatividade tem sido ligada ao sistema DA no caso da esquizofrenia (Eysenck, 1993). Embora vários estudos mostram que pacientes com esquizofrenia e personalidade esquizotípica exibem alta criatividade (O’Reilly et al., 2001; Folley e Park, 2005), modelos recentes da esquizofrenia e criatividade suporte de um U invertido em forma de associação em que a criatividade pode ser maior com o baixo-moderado transtorno esquizotípico de traço, mas diminui à medida que a gravidade da psicopatologia na esquizofrenia aumenta (Tsakanikos e Claridge, 2005; Stoneham e Coughtrey, 2009; Nelson e Rawlings, 2010). De facto, num estudo recente, Jaracz et al. (2012) relatou que os pacientes com esquizofrenia mostram criatividade diminuída em comparação com os controles. Além disso, os autores relataram que a criatividade comprometida estava associada com baixa flexibilidade cognitiva e impulsividade, refletida por suas baixas pontuações no teste de classificação de cartões de Wisconsin (WCST). Estes resultados estão em linha com pesquisas anteriores mostrando que os pacientes com esquizofrenia crônica exibem menor criatividade, que no caso de fluência criativa foi mediada por pontuações em tarefas de controle executivo e no caso de originalidade não foi (Abraham et al., 2007). Coletivamente, enquanto a impulsividade funcional leve pode melhorar a criatividade entre pacientes com características esquizotípicas, especialmente na geração de ideias que diferem dos exemplos dados (Abraham e Windmann, 2008), a impulsividade disfuncional observada na esquizofrenia grave pode diminuir as habilidades criativas.
os resultados do estudo actual apoiam a relação entre a criatividade e as variantes dos polimorfismos genéticos (Reuter et al., 2006; Runco et al., 2011). Além disso, os resultados encontrados em nossa amostra indicam que o principal efeito do alelo DRD4-7R na criatividade pode ser devido ao seu impacto na flexibilidade. Vários estudos associaram a flexibilidade cognitiva às vias DA (Ashby e Isen, 1999; Dreisbach e Goschke, 2004; Müller et al., 2007). Tem sido postulado que a ativação de receptores do tipo D2 (ou seja, D2, D3 e D4) diminui a inibição GABAérgica e facilita a ativação de múltiplas representações, aumentando assim a flexibilidade manifestada por habilidades de mudança de conjuntos (Müller et al., 2007; Seamans and Robbins, 2010). Além disso, os receptores do tipo D2 estão ligados à integração flexível de novas informações (Durstewitz e Seamans, 2008). Em linha com isto, Reuter et al. (2006) encontrou uma associação entre o receptor de dopamina D2 e a criatividade, medida por um índice composto, embora não seja possível determinar se a flexibilidade desempenhou um papel importante nestes resultados. Além disso, Runco et al. (2011) relatou diferenças na fluência criativa entre portadores de vários genes dopaminérgicos, entre eles o DRD4. No entanto, não está claro no seu relatório que grupo exibe maiores capacidades criativas. É importante notar que, ao contrário dos resultados aqui apresentados, Runco et al. (2011) não encontrou uma associação entre as medidas de flexibilidade medidas medidas pelas tarefas DT e DRD4. Esta inconsistência pode ser explicada pelas diferenças nas populações utilizadas e nas dimensões dos grupos ou pelas diferenças nas frequências dos alelos na população amostrada. Além disso, existem evidências sugerindo que a associação entre a dopamina e a DT pode não ser linear (Akbari Chermahini e Hommel, 2010, 2012), indicando que a interação entre vários DA genes podem influenciar os resultados individuais de cada variabilidade genética no contexto do DT. Além disso, vários estudos têm apontado o fato de que as vias Da podem ser sensíveis ao gênero (Tammimäki e Männistö, 2011; Wang et al., 2012). Zhong et al. (2010), por exemplo, descobriu que o gênero modulava a associação entre o polimorfismo DRD4 exon3 e o senso de Justiça. No presente estudo não encontramos um efeito de interação entre o DRD4-7R e de gênero, indicando que o gênero não tem um efeito modulador sobre a associação entre DRD4 e criatividade em nossa amostra. Além disso, os resultados aqui apresentados indicam uma falta de diferenças de género na criatividade e, em particular, na DT (para revisão, ver Baer e Kaufman, 2008).em conclusão, no presente estudo sugerimos que a associação encontrada entre DRD4 e a criatividade é principalmente influenciada pela flexibilidade. Os resultados do presente estudo estão em linha com os resultados anteriores apontando para o envolvimento do sistema DA criatividade e adicionar o conjunto de conhecimentos acumulados, sugerindo influências genéticas na criatividade.existem várias limitações do estudo que devem ser reconhecidas. Em primeiro lugar, o presente estudo apenas examinou a associação entre uma variabilidade genética no sistema DA—O DRD4—e DT. No entanto, existem outros polimorfismos importantes no sistema estriatal DA que podem ser associados com DT e podem contribuir tanto de forma autônoma como em interações gene-gene. Um desses polimorfismos é o transportador de dopamina (DAT1), que demonstrou estar associado à inibição do retorno (Colzato et al., 2010), bem como com medidas de impulsividade (Gizer e Waldman, 2012). Além disso, como mencionado acima, outra possibilidade que não foi explorada aqui é os possíveis efeitos de interação gene-gene sobre DT. Akbari Chermahini e Hommel (2010) relataram não-linear associação entre um marcador de striatal dopamina e DT, indicando que a relação pode ser mais complexo, de tal modo que um polimorfismo do gene pode afetar DT através da mediação de um outro gene polimorfismo. Assim, estudos futuros devem considerar examinar a interação entre estes dois polimorfismos no contexto da flexibilidade e DT.
Declaração de conflito de interesses
os autores declaram que a investigação foi realizada na ausência de quaisquer relações comerciais ou financeiras que possam ser interpretadas como um potencial conflito de interesses.Akbari Chermahini, S., and Hommel, B. (2010). A ligação entre a criatividade e a dopamina: as taxas espontâneas de piscar de olhos prevêem e dissociam o pensamento divergente e convergente. Cognition 115, 458-465. doi: 10.1016/j. cognition.2010.03.007
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Cools, R. (2008). Papel da dopamina no controle motivacional e cognitivo do comportamento. Neurocientista 14, 381-395. doi: 10.1177 / 1073858408317009 Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Frank, M. J. (2005). Modulação dinâmica da dopamina nos gânglios basais: a neurocomputational account of cognitive deficits in medicated and não medicated Parkinsonism. J. Cogn. Neuroci. 17, 51–72. doi: 10.1162 / 0898929052880093
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Healey, D., and Rucklidge, J. J. (2006). Uma investigação sobre a relação entre sintomatologia de TDAH, criatividade e funcionamento neuropsicológico em crianças. Neuropsicol Infantil. 12, 421–438. doi: 10.1080/09297040600806086
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Runco, M. A., and Richards, R. (1997). Criatividade eminente, criatividade cotidiana e Saúde. Greenwich, CT: Ablex.
Torrance, E. (1974). As diferenças não são défices. Ensinar. Coll. Gravacao. 75, 471–487.