- Introducere
- metoda
- participanți
- evaluarea creativității
- testul Torrance al gândirii Creative (cercuri Sub-scară)
- sarcina utilizări alternative (AUT)
- Pretest
- extracția și genotiparea ADN-ului
- analiza statistică
- rezultate
- componenta de fluență
- componenta de flexibilitate
- componenta de originalitate
- diferențe de gen
- discuție
- Declarație privind conflictul de interese
Introducere
cunoașterea creativă joacă un rol important în artă, în invenție și inovație, precum și în viața de zi cu zi (Runco și Richards, 1997). Deși creativitatea a fost considerată o capacitate umană unică, comportamentele Creative spontane (de ex., crearea de noi instrumente printre primate și păsări) s-au dovedit a apărea și la non-oameni (Byrne și Bates, 2007), atestând în continuare rădăcinile evolutive și biologice profunde ale creativității. Creativitatea a fost definită ca abilitatea de a produce răspunsuri care sunt atât noi (adică originale, rare și neașteptate), cât și potrivite (adică adaptabile și utile în funcție de constrângerile de sarcini) (Sternberg și Lubart, 1999). Una dintre abordările psihometrice ale măsurării creativității implică sarcini de gândire divergentă (DT) în care participanții sunt rugați să răspundă la o problemă dată cu soluții multiple (Dietrich și Kanso, 2010). Testele DT măsoară în general diferite aspecte ale creativității, inclusiv fluența creativă, flexibilitatea și originalitatea (Torrance, 1974). Astfel, deși nu sunt sinonime cu creativitatea, sarcinile DT oferă măsurători structurate și obiective ale creativității (Sternberg și Lubart, 1999; Jung și colab., 2009). În special, scorurile privind sarcinile DT s-au dovedit a fi corelate pozitiv cu măsurile de realizare creativă valabile din punct de vedere ecologic (Carson și colab., 2005), precum și cu creativitate auto-evaluată (Furnham și Bachtiar, 2008).
studiile anterioare au indicat implicarea sistemului dopaminergic (DA) în creativitate (Heilman și colab., 2003; Flaherty, 2005; Takeuchi și colab., 2010). Takeuchi și colab. (2010) a constatat că diferențele individuale în creativitate, măsurate prin DT, sunt corelate pozitiv cu materia cenușie în regiunile sistemului DA, inclusiv cortexul prefrontal dorsolateral, ganglionii bazali bilaterali, substantia nigra și zona ventrală tegmentală. În plus, mai multe studii genetice au arătat o relație între DT și neurotransmisia dopaminei (Reuter și colab., 2006; de Manzano și colab., 2010; Runco și colab., 2011). De exemplu, Reuter și colab. (2006) a constatat că creativitatea, măsurată prin SARCINI DT care implică atât creativitatea figurală, cât și cea verbală, este asociată semnificativ cu polimorfismele genei receptorului dopaminei D2 (DRD2). Mai mult, mai multe studii au descoperit că dopamina corticală este implicată în flexibilitatea cognitivă (Frank, 2005; Cools, 2008; Garcia-Garcia și colab., 2010), una dintre componentele principale ale DT.
o genă candidată DA interesantă pentru creativitate este gena receptorului dopaminei D4 (DRD4). Receptorul DRD4 este unul dintre cei cinci receptori ai dopaminei și joacă un rol important în medierea semnalizării dopaminei sinaptice. Gena se caracterizează printr-un număr variabil de 48 de perechi de baze de repetări tandem (48-bp VNTR) situate în regiunea de codificare a celui de-al treilea exon. DRD4 48-BP polimorfismul VNTR variază de la 2 la 11 repetări la indivizi (Asghari și colab., 1995; Cravchik și Goldman, 2000), unde repetarea 4 (4R) este cea mai frecventă repetare la populațiile caucaziene, iar repetarea 7 (7R) este a doua variantă cea mai comună (Chang și colab., 1996; Ding și colab., 2002). Interesant este că 7R a fost asociat anterior cu comportamente creative din viața reală, cum ar fi trăsătura de personalitate care caută noutate (Ebstein și colab., 1996). Într-adevăr, căutarea noutății—tendința spre activitatea exploratorie—este considerată a fi una dintre caracteristicile oamenilor creativi (Chavez-Eakle și colab., 2006; Drago și colab., 2009). Deși dovezile pentru o asociere între DRD4 7R și căutarea noutății sunt inconsistente, după cum reiese din mai multe rapoarte de meta-analiză (Kluger și colab., 2002; Munaf XV și colab., 2008), multe studii au găsit o asociere semnificativă între căutarea noutății și alela 7R a DRD4 (Ebstein și colab., 1996; Benjamin și colab., 2000; Keltikangas-J Oustrvinen și colab., 2002; Becker și colab., 2005). Roussos și colab. (2009) au sugerat recent că diferențele în măsurarea căutării noutății folosind scale de auto-raportare pot explica rezultatele inconsistente dintre polimorfismul 7R și căutarea noutății. În conformitate cu aceasta, s-a constatat în mod repetat că sistemul DA joacă un rol major în trăsătura de personalitate a căutării noutății (Flaherty, 2005; Schweizer, 2006), atestând în continuare rolul potențial al DRD4 în creativitate.cu toate acestea, spre deosebire de rolul DRD4 48-bp VNTR 7R în căutarea noutății, dovezile recente indică faptul că 7R este de fapt asociat cu o flexibilitate cognitivă afectată, unul dintre aspectele majore ale creativității (Strobel și colab., 2004; Congdon și colab., 2008). Flexibilitatea implică capacitatea de a face un răspuns alternativ după inhibarea cu succes a unui răspuns curent. Constatările de până acum privind asocierea dintre inhibiție și DRD4 sunt mixte. Congdon și colab. (2008) a constatat că participanții cu alela 7R a DRD4 prezintă un timp de reacție mai mare a semnalului de oprire (SSRT) într-o sarcină go/no-go, reflectând un control inhibitor mai slab, în timp ce Forbes și colab. (2009) nu a reușit să găsească un astfel de efect asupra Scala impulsivității Barratt (BIS), un instrument de auto-raportare pentru măsurarea impulsivității. Pe de altă parte, Colzato și colab. (2010) a demonstrat că aceste efecte mixte se pot datora faptului că studiile anterioare au abordat impulsivitatea ca proces monolitic, în timp ce impulsivitatea poate fi de fapt împărțită în tipuri funcționale și disfuncționale conform inventarului impulsivității lui Dickman (DII; Dickman, 1990). Impulsivitatea disfuncțională este tendința de a acționa fără preconcepție în cazurile în care o astfel de acțiune este inadecvată, în timp ce impulsivitatea funcțională este o tendință similară implementată în situații adecvate (Colzato și colab., 2010). Colzato și colab. (2010) a constatat că persoanele cu DRD4 7R, care este asociat cu niveluri mai ridicate de da striatală, au prezentat impulsivitate disfuncțională mai mare. Mai multe studii meta-analitice au indicat alela 7R a DRD4 ca o alelă de risc pentru tulburarea de hiperactivitate cu deficit de atenție (ADHD; Faraone și colab., 2001; DiMaio și colab., 2003), o tulburare caracterizată prin impulsivitate disfuncțională ridicată (Young și colab., 2007). Mai mult, relația dintre ADHD și creativitate a fost investigată în mai multe studii (Shaw și Brown, 1990; Healey și Rucklidge, 2006), deși până acum aceste studii au dat rezultate neclare. În timp ce unii au descoperit că ADHD și simptomologia sa sunt corelate pozitiv cu creativitatea (Healey și Rucklidge, 2006; White și Shah, 2011), alții au raportat o tendință opusă (Funk și colab., 1993; Healey și Rucklidge, 2005, 2008). Interesant este că ADHD este asociat comportamental cu deficiențe în controlul executiv, inclusiv schimbarea seturilor atenționale (Boonstra și colab., 2005) și flexibilitate (Barkley și colab., 1997; sergent și colab., 2002), ambele fiind importante pentru flexibilitatea cognitivă, o dimensiune centrală a creativității (Dietrich, 2004; Durstewitz și Seamans, 2008). Aceste constatări indică faptul că persoanele cu alela 7R a DRD4 pot prezenta niveluri mai scăzute de creativitate și flexibilitate diminuată în special în ceea ce privește sarcinile DT. Prin urmare, DRD4 exon III VNTR este un candidat biologic plauzibil pentru a contribui la diferențele individuale în creativitate.
în mod colectiv, se pare că, pe de o parte, alela 7R, ca alelă de risc pentru funcțiile executive afectate, flexibilitatea și schimbarea setului, poate fi asociată cu o creativitate scăzută. Pe de altă parte, asocierea dintre alela 7R și căutarea noutății superioare indică faptul că indivizii cu alela 7R pot prezenta de fapt o mai mare flexibilitate și creativitate.
studiul actual a fost conceput pentru a explora rolul complex al DRD4 7R în modelarea parțială a creativității umane. Pentru a caracteriza mai bine această relație, am evaluat două tipuri de teste DT (figural și verbal). Punctajul fiecărei sarcini a fost împărțit în funcție de cele trei dimensiuni principale ale DT, și anume fluența creativă, flexibilitatea și originalitatea și fiecare dimensiune a fost analizată separat pentru a testa ipoteza că DRD4 afectează diferențiat fiecare dimensiune a creativității.
metoda
participanți
eșantionul a cuprins 185 de studenți de la Universitatea din Haifa din Israel (112 femei). Toți participanții au fost caucazieni (auto-raportați), cu o vârstă medie de 24, 5 ani (DS 2, 1). Participanții au fost recrutați prin anunțuri postate la universitate. Toți participanții au fost voluntari plătiți.
evaluarea creativității
creativitatea a fost evaluată folosind două teste DT, o sub-scară din sub-testul figural al testului Torrance al gândirii creative (Torrance, 1974) și sarcina utilizări alternative (aut; Guilford și colab., 1978). Ambele teste au inclus cele trei dimensiuni de bază ale DT, și anume flexibilitatea, originalitatea și fluența.
testul Torrance al gândirii Creative (cercuri Sub-scară)
participanților li s-a oferit o pagină pe care au fost desenate 30 de cercuri identice. Li s-a cerut să deseneze cât mai multe obiecte semnificative într-un termen de 10 minute, unde fiecare desen trebuie să includă cel puțin un cerc. Punctajul a inclus fluența (numărul de răspunsuri produse), flexibilitatea (numărul de categorii) și originalitatea, calculate în funcție de punctajul răspunsurilor originale, așa cum este detaliat în Ghidul de notare Torrance Tests of Creative Thinking (Torrance, 1974).
sarcina utilizări alternative (AUT)
participanților li s-a oferit o listă cu cinci obiecte comune și li s-a cerut să enumere cât mai multe utilizări alternative pentru fiecare obiect, într-un termen de 10 minute. Cea mai obișnuită utilizare zilnică a fost indicată în paranteze. Obiectele au fost: pantof (uz comun: uzura pe jos); buton (uz comun: lucruri de închidere); Creion (uz comun: desen sau scris); anvelope (uz comun: roata auto); și pahar (uz comun: conține lichid). Numai răspunsurile care nu au reiterat utilizările comune date au fost numărate și incluse. Ca și în testul Torrance, punctajul a inclus fluență, flexibilitate și originalitate. Deoarece nu există linii directoare pentru punctarea răspunsurilor originale în AUT, răspunsurile originale au fost definite ca răspunsuri statistice rare, conform unui pretest efectuat în laboratorul nostru.
Pretest
în scopul creării unui criteriu valid al frecvenței de răspuns, un grup de 100 de participanți sănătoși care nu au participat la acest studiu au finalizat AUT. Pentru fiecare obiect, a fost colectată o listă cu toate utilizările posibile de la toți participanții. O măsură statistică a frecvenței a fost calculată pe baza acestei liste pentru a evalua scorul de originalitate pentru fiecare răspuns și, ulterior, pentru fiecare participant. Răspunsurilor li s-a atribuit un scor de zero dacă 5% sau mai mulți dintre participanți au enumerat o utilizare dată, un scor de unul dacă între 2% și 4,99% dintre participanți l-au enumerat și un scor de două dacă mai puțin de 1,99% au enumerat utilizarea. Un scor mediu de originalitate a fost calculat pentru fiecare participant în funcție de aceste scoruri statistice rare.
extracția și genotiparea ADN-ului
ADN-ul a fost extras din 20 ml de probe de apă de gură folosind kitul Master Pure (Epicentre, Madison, WI). DRD4 48-bp VNTR a fost caracterizat printr – o procedură de amplificare PCR cu următorii primeri: F5′ – CTT CCT ACC CTG CCC GCT CAT GCT GCTGCT CTA CTGG – 3′ și R5′ – ACC ACCACC GGC AGG ACC CTC ATG GCC TTG CGC TC-3. Reacțiile PCR s-au realizat folosind 5 unktil Master Mix (Thermo scientific), 2 unktil primeri (0,5 unktilm), 0,6 oktil mg/Cl2 (2,5 mM), 0.În amestec s-au adăugat 4 0tql DMSO 5% și 1 xtql de apă la un volum total de 9 xtql și încă 1 ztql de ADN genomic. Toate reacțiile PCR au fost efectuate pe un Termocicler Biometra T1 (Biometra, G Inkttingem, Germania). Starea de reacție PCR a fost următoarea: etapa de preîncălzire la 94,0 CT pentru 5 min, 34 cicluri de denaturare la 94,0 CT pentru 30 s, reanalizarea la 55 CT pentru 30 s și extinderea la 72 CT pentru 90 s. reacția s-a menținut la 72 CT pentru 5 min. Amestecul de reacție a fost apoi electroforezat pe un gel de agaroză 3% (AMRESCO) cu bromură de etidiu pentru a detecta genotipurile.
distribuția frecvenței genotipului a fost în funcție de echilibrul Hardy-Weinberg (chi-pătrat = 0,01, valoarea p = 0,92).
analiza statistică
genotipul a fost clasificat în funcție de prezența sau absența alelei 7R a DRD4 (7 vs.no7), ca în conformitate cu rapoartele anterioare din literatură (Ding și colab., 2002). Diferențele de grup privind scorurile de creativitate au fost analizate statistic folosind ANOVAs, cu prezența (7) sau absența (no7) a alelei DRD4 7R ca variabilă independentă. Pentru a examina diferitele aspecte ale DT (fluență, flexibilitate, originalitate), am efectuat o analiză multivariată a varianței (MANOVA) separat pentru fiecare componentă.
rezultate
pentru a confirma că cele două grupuri (7, no7) nu au fost semnificativ diferite în ceea ce privește vârsta și educația, am efectuat teste t independente pentru a compara cele două grupuri (vezi Tabelul 1 Pentru mijloace și abateri standard). Această analiză a arătat că cele două grupuri nu diferă în ceea ce privește vârsta sau anii de educație. Testele non-parametrice (Mann-Whitney) au evidențiat diferențe semnificative în distribuția frecvenței pentru sex (Z = 2.11, p < 0.05).
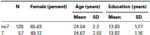
Tabelul 1. Date privind vârsta, sexul și educația pentru genotipul DRD4 7R, SD: deviație standard.
scorurile medii ale măsurilor de creativitate utilizate în cele două sarcini sunt rezumate în tabelul 2.

Tabelul 2. Mijloace și eroare Standard (SE) a celor două sarcini de creativitate utilizate: AUT și Torrance.
componenta de fluență
un ANOVA multivariat a indicat un efect general al genotipului DRD4 7R pentru componenta de fluență, F(2,182) = 3,25, p <0,05. Având în vedere semnificația testului global, au fost examinate efectele principale univariate. Principalele efecte univariate semnificative pentru genotipul DRD4 au fost obținute pentru fluența AUT F (1,183)=4,95, p < 0.05 (alela 7R care prezintă o fluență mai mică comparativ cu lotul no7, figura 1a), dar nu a atins nivelul de semnificație pentru fluența Torrance F(1,183) = 2,95, p = 0,087.
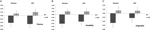
Figura 1. Scorurile creativității pe sarcinile AUT și Torrance separate de fluență (a), flexibilitate (B) și originalitate (C). Mijloacele au fost transformate în scoruri Z.
componenta de flexibilitate
Anova multivariată a măsurilor de flexibilitate a prezentat un efect general pentru DRD4 7R genotipul F(2,182) = 4.38, p < 0.05. Efecte principale univariate semnificative pentru genotipul DRD4 au fost obținute pentru flexibilitatea AUT F(1183)=4,57, p < 0,05 și pentru flexibilitatea Torrance F (1183)=6,95, p < 0,01. După cum se arată în figura 1B, în ambele cazuri, indivizii din grupul de alele 7R au prezentat o flexibilitate mai mică comparativ cu indivizii din grupul no7 (Tabelul 2).
componenta de originalitate
Anova multivariată a măsurilor de originalitate nu a prezentat efecte de semnificație, F(2,182) = 2,06, p = 0.13, deși scorurile au indicat o originalitate mai mică pentru alela 7R a DRD4 (a se vedea figura 1C, Tabelul 2 Pentru detalii).
diferențe de gen
discuție
sarcinile DT necesită participanților să ofere soluții multiple la o anumită problemă. În sarcinile utilizate aici, participanților li s-a cerut să se gândească la multe utilizări alternative posibile pentru obiectele de zi cu zi (în AUT) și să deseneze mai multe desene care încorporează o formă specifică (cerc) (în sarcina Torrance). Rezultatele prezentate aici demonstrează că indivizii cu alela DRD4 7R prezintă o creativitate mai mică măsurată prin DT. Având în vedere că alela 7R a fost asociată cu impulsivitate disfuncțională și inhibiție slabă (Congdon și colab., 2008; Dreber și colab., 2009), faptul că indivizii cu alela 7R prezintă o creativitate mai mică poate fi explicat prin incapacitatea lor de a suprima sau inhiba răspunsurile evidente (comune), după cum indică scorurile de originalitate mai mici. Scorurile mai mici de flexibilitate și fluență pot indica faptul că, odată ce a fost dat un răspuns comun, devine mai greu să treci la o nouă categorie (așa cum este indicat de scorurile scăzute de flexibilitate), ducând la mai puține răspunsuri (așa cum este indicat de scorul scăzut de fluență). Dovezi suplimentare pentru relația dintre creativitate și impulsivitate pot fi văzute în asocierea alelei 7R cu ADHD (Li și colab., 2006), precum și cu impulsivitate crescută la pacienții cu ADHD (Langley și colab., 2004). Interesant este că creativitatea a fost legată de sistemul DA În cazul schizofreniei (Eysenck, 1993). Deși mai multe studii arată că pacienții cu schizofrenie și personalitate schizotipală prezintă o creativitate ridicată (O ‘ Reilly și colab., 2001; Folley și Park, 2005), modelele recente de schizofrenie și creativitate susțin o asociere inversată în formă de U în care creativitatea poate fi mai mare cu trăsătură schizotipală scăzută-moderată, dar scade pe măsură ce severitatea psihopatologiei în schizofrenie crește (Tsakanikos și Claridge, 2005; Stoneham și Coughtrey, 2009; Nelson și Rawlings, 2010). Într-adevăr, într-un studiu recent Jaracz și colab. (2012) a raportat că pacienții cu schizofrenie prezintă o creativitate diminuată în comparație cu controalele. Mai mult, autorii au raportat că creativitatea afectată a fost asociată cu flexibilitate cognitivă scăzută și impulsivitate, reflectată de scorurile lor scăzute la testul de sortare a cardurilor din Wisconsin (WCST). Aceste rezultate sunt în concordanță cu cercetările anterioare care arată că pacienții cu schizofrenie cronică prezintă o creativitate mai mică, care în cazul fluenței creative a fost mediată de scoruri privind sarcinile de control executiv și în cazul originalității nu a fost (Abraham și colab., 2007). În mod colectiv, în timp ce impulsivitatea funcțională ușoară poate îmbunătăți creativitatea în rândul pacienților cu trăsături schizotipale, în special în generarea de idei care diferă de exemplele date (Abraham și Windmann, 2008), impulsivitatea disfuncțională observată în schizofrenia severă poate diminua abilitățile creative.
rezultatele studiului actual susțin relația dintre creativitate și variantele polimorfismelor genetice DA (Reuter și colab., 2006; Runco și colab., 2011). Mai mult, rezultatele găsite în eșantionul nostru indică faptul că efectul principal al alelei DRD4-7R asupra creativității se poate datora impactului său asupra flexibilității. Mai multe studii au legat flexibilitatea cognitivă cu căile DA (Ashby și Isen, 1999; Dreisbach și Goschke, 2004; m Okticller și colab., 2007). S-a postulat că activarea receptorilor asemănători D2 (adică D2, D3 și D4) scade inhibarea GABAergică și facilitează activarea reprezentărilor multiple, crescând astfel flexibilitatea manifestată prin abilități de schimbare a seturilor (m Electicller și colab., 2007; Seamans și Robbins, 2010). Mai mult, receptorii asemănători D2 au fost legați de integrarea flexibilă a informațiilor noi (Durstewitz și Seamans, 2008). În conformitate cu aceasta, Reuter și colab. (2006) a găsit o asociere între receptorul dopaminei D2 și creativitate măsurată printr-un indice compozit, deși nu se poate determina dacă flexibilitatea a jucat un rol major în aceste rezultate. În plus, Runco și colab. (2011) au raportat diferențe în fluența creativă în rândul purtătorilor mai multor gene de dopamină, printre care DRD4. Cu toate acestea, din raportul lor nu este clar care grup prezintă abilități creative mai mari. Este important de menționat că, spre deosebire de rezultatele prezentate aici, Runco și colab. (2011) nu a găsit o asociere între măsurile de flexibilitate măsurate prin SARCINI DT și DRD4. Această inconsecvență poate fi explicată prin diferențe între populațiile utilizate și dimensiunile grupurilor sau prin diferențe între frecvențele alelelor din cadrul populației incluse în eșantion. Mai mult, există dovezi care sugerează că asocierea dintre dopamină și DT poate să nu fie una liniară (Akbari Chermahini și Hommel, 2010, 2012), indicând faptul că o interacțiune între mai multe gene DA poate influența rezultatele individuale ale fiecărei variabilități genetice în contextul DT. În plus, mai multe studii au subliniat faptul că căile DA pot fi sensibile la Gen (Tammim Oktokki și m Oktoknnist Oktokt, 2011; Wang și colab., 2012). Zhong și colab. (2010), de exemplu, a constatat că genul a modulat asocierea dintre polimorfismul DRD4 exon3 și simțul corectitudinii. În studiul actual nu am găsit un efect de interacțiune între DRD4-7R și gen, indicând faptul că genul nu are un efect modulator asupra asocierii dintre DRD4 și creativitate în eșantionul nostru. În plus, rezultatele prezentate aici indică o lipsă a diferențelor de gen în creativitate și în special în DT (pentru revizuire, a se vedea Baer și Kaufman, 2008).
În concluzie, în studiul actual sugerăm că asocierea găsită între DRD4 și creativitate este influențată în principal de flexibilitate. Rezultatele studiului actual sunt în concordanță cu rezultatele anterioare care indică implicarea sistemului DA în creativitate și se adaugă corpului acumulat de cunoștințe care sugerează influențe genetice în creativitate.
există mai multe limitări ale studiului care ar trebui recunoscute. În primul rând, studiul actual a examinat doar asocierea dintre o variabilitate genetică în sistemul DA—DRD4—și DT. Cu toate acestea, există și alte polimorfisme importante în sistemul Da striatal care ar putea fi asociate cu DT și pot contribui atât într-o manieră independentă, cât și în interacțiunile genă-genă. Un astfel de polimorfism este transportorul de dopamină (DAT1), care s-a dovedit a fi asociat cu inhibarea revenirii (Colzato și colab., 2010), precum și cu măsuri de impulsivitate (Gizer și Waldman, 2012). Mai mult, după cum sa menționat mai sus, o altă posibilitate care nu a fost explorată aici este posibilele efecte de interacțiune genă-genă asupra DT. Akbari Chermahini și Hommel (2010) au raportat o asociere neliniară între un marker al dopaminei striatale și DT, indicând faptul că relația poate fi mai complexă, astfel încât un polimorfism al unei gene poate avea impact asupra DT prin medierea unui alt polimorfism genic. Astfel, studiile viitoare ar trebui să ia în considerare examinarea interacțiunii dintre aceste două polimorfisme în contextul flexibilității și DT.
Declarație privind conflictul de interese
autorii declară că cercetarea a fost realizată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.Akbari Chermahini, S. și Hommel, B. (2010). (B) legătura dintre creativitate și dopamină: ratele spontane de clipire a ochilor prezic și disociază gândirea divergentă și convergentă. Cunoașterea 115, 458-465. doi: 10.1016 / j. cunoaștere.2010.03.007
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Cools, R. (2008). Rolul dopaminei în controlul motivațional și cognitiv al comportamentului. Neurolog 14, 381-395. doi: 10.1177 / 1073858408317009
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Frank, M. J. (2005). Modularea dinamică a dopaminei în ganglionii bazali: un cont neurocomputațional al deficitelor cognitive în Parkinsonismul medicamentos și nemedicat. J. Cogn. Neuroști. 17, 51–72. doi: 10.1162 / 0898929052880093
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Healey, D. și Rucklidge, J. J. (2006). O investigație a relației dintre simptomatologia ADHD, creativitatea și funcționarea neuropsihologică la copii. Neuropsihol Pentru Copii. 12, 421–438. doi: 10.1080 / 09297040600806086
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Runco, M. A. și Richards, R. (1997). Creativitate eminentă, creativitate de zi cu zi și sănătate. Greenwich, CT: ABLEX.
Torrance, E. (1974). Diferențele nu sunt deficite. Preda. Col. Rec. 75, 471–487.