- introduktion
- metod
- deltagare
- bedömning av kreativitet
- Torrance Test of Creative Thinking (Circles Sub-Scale)
- Alternate Uses Task (Aut)
- Pretest
- DNA-extraktion och genotypning
- statistisk analys
- resultat
- Flytkomponenten
- Flexibilitetskomponenten
- Originalitetskomponenten
- könsskillnader
- diskussion
- intressekonflikt uttalande
introduktion
Kreativ kognition spelar en viktig roll i konsten, i uppfinning och innovation, såväl som i vardagen (Runco och Richards, 1997). Även om kreativitet har ansetts vara en unik mänsklig kapacitet, spontant kreativa beteenden (t. ex., skapa nya verktyg bland primater och fåglar) har visat sig förekomma hos icke-människor också (Byrne och Bates, 2007), vilket ytterligare bekräftar kreativitetens djupa evolutionära och biologiska rötter. Kreativitet har definierats som förmågan att producera svar som är både nya (dvs. original, sällsynta och oväntade) och lämpliga (dvs. adaptiva och användbara enligt uppgiftsbegränsningar) (Sternberg och Lubart, 1999). En av de psykometriska metoderna för att mäta kreativitet innebär divergerande tänkande (DT) uppgifter där deltagarna uppmanas att svara på ett givet problem med flera lösningar (Dietrich och Kanso, 2010). Tester av DT mäter i allmänhet olika aspekter av kreativitet, inklusive kreativ flyt, flexibilitet och originalitet (Torrance, 1974). Således, även om det inte är synonymt med kreativitet, ger DT-uppgifter strukturerade och objektiva mätningar av kreativitet (Sternberg och Lubart, 1999; Jung et al., 2009). I synnerhet har poäng på DT-uppgifter visat sig vara positivt korrelerade med ekologiskt giltiga mått på kreativ prestation (Carson et al., 2005) samt med självskattad kreativitet (Furnham och Bachtiar, 2008).
tidigare studier har pekat på involveringen av det dopaminerga (DA) systemet i kreativitet (Heilman et al., 2003; Flaherty, 2005; Takeuchi et al., 2010). Takeuchi et al. (2010) fann individuella skillnader i kreativitet, mätt med DT, att vara positivt korrelerade med grå substans i DA-systemregioner, inklusive den dorsolaterala prefrontala cortexen, bilaterala basala ganglier, substantia nigra och det tegmentala ventrala området. Dessutom har flera genetiska studier visat ett samband mellan dt och dopamin neurotransmission (Reuter et al., 2006; De Manzano et al., 2010; Runco et al., 2011). Till exempel, Reuter et al. (2006) fann kreativitet, mätt med DT-uppgifter som involverar både figural och verbal kreativitet, att vara signifikant associerad med polymorfismer av dopamin D2-receptorgenen (DRD2). Dessutom har flera studier funnit att kortikal dopamin är involverad i kognitiv flexibilitet (Frank, 2005; Cools, 2008; Garcia-Garcia et al., 2010), en av huvudkomponenterna i DT.
en intressant da-kandidatgen för kreativitet är dopamin D4-receptorgenen (DRD4). DRD4-receptorn är en av fem dopaminreceptorer och spelar en viktig roll för att förmedla synaptisk dopaminsignalering. Genen kännetecknas av ett 48-basparvariabelantal tandemupprepningar (48-bp VNTR) som ligger i kodningsområdet för den tredje exonen. DRD4 48-bp VNTR polymorfism varierar från 2 till 11 upprepningar mellan individer (Asghari et al., 1995; Cravchik och Goldman, 2000), där 4-upprepningen (4R) är den vanligaste upprepningen i kaukasiska populationer och 7-upprepningen (7R) är den näst vanligaste varianten (Chang et al., 1996; Ding et al., 2002). Intressant nog har 7R tidigare associerats med verkliga kreativa beteenden, såsom det nyhetssökande personlighetsdraget (Ebstein et al., 1996). Faktum är att nyhetssökande-tendensen till utforskande aktivitet—anses vara en av egenskaperna hos kreativa människor (Chavez-Eakle et al., 2006; Drago et al., 2009). Även om bevisen för en koppling mellan DRD4 7R och nyhetssökande är inkonsekvent, vilket framgår av flera metaanalysrapporter (Kluger et al., 2002; Munaf Ukrainian et al., 2008) har många studier funnit en signifikant koppling mellan nyhetssökande och 7R-allelen i DRD4 (Ebstein et al., 1996; Benjamin et al., 2000; Keltikangas-J., 2002; Becker et al., 2005). Roussos et al. (2009) har nyligen föreslagit att skillnader i mätning av nyhetssökande med hjälp av självrapportskalor kan redogöra för de inkonsekventa resultaten mellan 7R-polymorfism och nyhetssökande. I linje med detta har det upprepade gånger visat sig att DA-systemet spelar en viktig roll i personlighetsdraget för nyhetssökande (Flaherty, 2005; Schweizer, 2006), vilket ytterligare bekräftar DRD4: s potentiella roll i kreativitet.
i motsats till rollen som DRD4 48-bp VNTR 7R i nyhetssökande, tyder nya bevis på att 7R faktiskt är förknippat med nedsatt kognitiv flexibilitet, en av de viktigaste aspekterna av kreativitet (Strobel et al., 2004; Congdon et al., 2008). Flexibilitet innebär förmågan att göra ett alternativt svar efter att ha lyckats hämma ett aktuellt svar. Fynd hittills om sambandet mellan hämning och DRD4 blandas. Congdon et al. (2008) fann att deltagare med 7R-allelen i DRD4 uppvisar högre stopp-signalreaktionstid (SSRT) på en go/no-go-uppgift, vilket återspeglar sämre hämmande kontroll, medan Forbes et al. (2009) misslyckades med att hitta en sådan effekt på Barratt Impulsiveness Scale (BIS), ett självrapporteringsverktyg för att mäta impulsivitet. Å andra sidan, Colzato et al. (2010) visade att dessa blandade effekter kan bero på det faktum att tidigare studier behandlade impulsivitet som en monolitisk process, medan impulsivitet faktiskt kan delas in i funktionella och dysfunktionella typer enligt Dickmans Impulsivitetsinventering (DII; Dickman, 1990). Dysfunktionell impulsivitet är tendensen att agera utan eftertanke i fall då sådan åtgärd är olämplig, medan funktionell impulsivitet är en liknande tendens implementerad i lämpliga situationer (Colzato et al., 2010). Colzato et al. (2010) fann att individer med DRD4 7R, som är associerad med högre nivåer av striatal DA, uppvisade högre dysfunktionell impulsivitet. Flera metaanalytiska studier har pekat på 7R-allelen i DRD4 som en riskallel för hyperaktivitetsstörning (ADHD; Faraone et al., 2001; DiMaio et al., 2003), en störning som kännetecknas av hög dysfunktionell impulsivitet (Young et al., 2007). Dessutom har förhållandet mellan ADHD och kreativitet undersökts i flera studier (Shaw och Brown, 1990; Healey och Rucklidge, 2006), men hittills har dessa studier gett oklara resultat. Medan vissa har funnit att ADHD och dess symptomologi är positivt korrelerade med kreativitet (Healey och Rucklidge, 2006; White and Shah, 2011), har andra rapporterat en motsatt trend (Funk et al., 1993; Healey och Rucklidge, 2005, 2008). Intressant är ADHD beteendemässigt förknippat med försämringar i verkställande kontroll, inklusive attentional set-shifting (Boonstra et al., 2005), och flexibilitet (Barkley et al., 1997; Sergeant et al., 2002), som båda är viktiga för kognitiv flexibilitet, en central dimension av kreativitet (Dietrich, 2004; Durstewitz och Seamans, 2008). Dessa resultat indikerar att individer med 7R-allelen i DRD4 kan uppvisa lägre nivåer av kreativitet och minskad flexibilitet på DT-uppgifter i synnerhet. Därför är DRD4 exon III VNTR en biologiskt trolig kandidat för att bidra till individuella skillnader i kreativitet.
sammantaget verkar det som att å ena sidan 7R-allelen, som en riskallel för nedsatt verkställande funktioner, flexibilitet och uppsättningsförskjutning, kan vara förknippad med låg kreativitet. Å andra sidan indikerar sambandet mellan 7R-allelen och högre nyhetssökande att individer med 7R-allelen faktiskt kan uppvisa större flexibilitet och kreativitet.
den aktuella studien har utformats för att undersöka den komplexa Roll DRD4 7R delvis forma mänsklig kreativitet. För att bättre karakterisera detta förhållande bedömde vi två typer av DT-test (figural och verbal). Poängen för varje uppgift delades upp enligt de tre huvuddimensionerna för DT, nämligen kreativ flyt, flexibilitet och originalitet, och varje dimension analyserades separat för att testa hypotesen att DRD4 differentiellt påverkar varje dimension av kreativitet.
metod
deltagare
provet omfattade 185 studenter från University of Haifa i Israel (112 kvinnor). Alla deltagare var kaukasiska (självrapporterade), med en medelålder på 24,5 (SD 2,1). Deltagarna rekryterades genom meddelanden som publicerades vid universitetet. Alla deltagare var betalda volontärer.
bedömning av kreativitet
kreativitet bedömdes med hjälp av två dt-tester, en delskala från figural sub-test av Torrance test of creative thinking (Torrance, 1974) och den alternativa användningen uppgift (Aut; Guilford et al., 1978). Båda testerna inkluderade de tre kärndimensionerna av DT, nämligen flexibilitet, originalitet och flyt.
Torrance Test of Creative Thinking (Circles Sub-Scale)
deltagarna fick en sida där 30 identiska cirklar ritades. De ombads att rita så många olika meningsfulla objekt som möjligt inom en tidsgräns på 10 min, där varje ritning måste innehålla minst en cirkel. Scoring inkluderade flyt (antal producerade svar), flexibilitet (antal kategorier) och originalitet, beräknat enligt poängen för ursprungliga svar, som beskrivs i Torrance Tests of Creative Thinking scoring guide (Torrance, 1974).
Alternate Uses Task (Aut)
deltagarna fick en lista med fem vanliga objekt och ombads att lista så många alternativa användningsområden som möjligt för varje objekt inom en tidsgräns på 10 minuter. Den vanligaste dagliga användningen indikerades inom parentes. Föremålen var: sko (vanligt bruk: slitage till fots); knapp (vanligt bruk: stängande saker); penna (vanligt bruk: ritning eller skrivning); däck (vanligt bruk: bilhjul); och dricksglas (vanligt bruk: innehåller vätska). Endast svar som inte upprepade de givna vanliga användningarna räknades och inkluderades. Som i Torrance-testet inkluderade poäng flytande, flexibilitet och originalitet. Eftersom det inte finns några riktlinjer för poängsättning av ursprungliga svar i AUT definierades ursprungliga svar som statistiskt sällsynta svar enligt ett förtest som genomfördes i vårt laboratorium.
Pretest
för att skapa ett giltigt kriterium för svarsfrekvens slutförde en grupp på 100 friska deltagare som inte deltog i denna studie AUT. För varje objekt samlades en lista över alla möjliga användningsområden från alla deltagare. Ett statistiskt sällan mått beräknades baserat på denna lista för att utvärdera originalitetspoängen för varje svar och därefter för varje deltagare. Svaren tilldelades en poäng på noll om 5% eller mer av deltagarna listade en given användning, en poäng på en om mellan 2% och 4,99% av deltagarna listade den och en poäng på två om mindre än 1,99% listade användningen. En genomsnittlig originalitetspoäng beräknades för varje deltagare enligt dessa statistiska sällsynta poäng.
DNA-extraktion och genotypning
DNA extraherades från 20 ml munvattenprover med hjälp av Master Pure kit (Epicentre, Madison, WI). DRD4 48-bp VNTR kännetecknades av en PCR – amplifieringsprocedur med följande primers: F5′ – CTT CCT ACC CTG CCC GCT CAT GCT GCTGCT CTA CTGG – 3′ och R5′ – ACC ACCACC GGC AGG ACC CTC ATG GCC TTG CGC TC-3. PCR-reaktioner utfördes med användning av 5 occl Master Mix (Thermo scientific), 2 occl primers (0,5 occlm), 0,6 occl Mg/Cl2 (2,5 mM), 0.4 oc 5% och 1 oc vatten till totalt 9 oc total volym och ytterligare 1 oc genomiskt DNA tillsattes till blandningen. Alla PCR-reaktioner utfördes på en Biometra T1-Termocyklist (Biometra, G. PCR-reaktionsförhållandet var följande: förvärmningssteg vid 94,0 C i 5 min, 34 cykler av denaturering vid 94,0 C i 30 s,reannealing vid 55 C i 30 s och förlängning vid 72 C i 90 s. reaktionen fortsatte att hålla vid 72 C i 5 min. Reaktionsblandningen elektrofores sedan på en 3% agarosgel (AMRESCO) med etidiumbromid för att screena för genotyper.
fördelningen av genotypfrekvensen var enligt Hardy-Weinberg-jämvikten (chi-kvadrat = 0,01, p-värde = 0,92).
statistisk analys
genotyp klassificerades enligt närvaron eller frånvaron av 7R-allelen i DRD4 (7 vs. no7), som i linje med tidigare rapporter i litteraturen (Ding et al., 2002). Gruppskillnader på kreativitetspoäng analyserades statistiskt med ANOVAs, med närvaron (7) eller frånvaron (no7) av DRD4 7R-allelen som den oberoende variabeln. För att undersöka de olika aspekterna av DT (flyt, flexibilitet, originalitet) genomförde vi en multivariat variansanalys (MANOVA) separat för varje komponent.
resultat
för att bekräfta att de två grupperna (7, no7) inte var signifikant olika vad gäller ålder och utbildning, genomförde vi oberoende t-test för att jämföra de två grupperna (se Tabell 1 för medel och standardavvikelser). Denna analys avslöjade att de två grupperna inte skilde sig åt när det gäller ålder eller utbildningsår. Icke-parametriska tester (Mann–Whitney) avslöjade signifikanta skillnader i frekvensfördelningen för kön (Z = 2.11, p < 0.05).
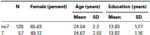
tabell 1. Ålder, kön och utbildningsdata för DRD4 7R genotyp, SD: standardavvikelse.
medelvärdena för kreativitetsmåtten som används i de två uppgifterna sammanfattas i Tabell 2.

tabell 2. Medel och standardfel (SE) av de två kreativitetsuppgifterna som används: AUT och Torrance.
Flytkomponenten
en multivariat ANOVA indikerade en allmän effekt av DRD4 7R-genotyp för flytkomponenten, F(2,182) = 3,25, p < 0,05. Med tanke på betydelsen av det övergripande testet undersöktes univariata huvudeffekter. Signifikanta univariata huvudeffekter för DRD4 genotyp erhölls för aut flyt F (1,183) = 4,95, p < 0.05 (7R-allel som uppvisar lägre flyt jämfört med no7-gruppen, Figur 1a), men nådde inte signifikansnivån för Torrance flyt F(1,183) = 2.95, p = 0.087.
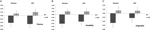
Figur 1. Kreativitet poäng på aut och Torrance uppgifter separerade till flyt (a), flexibilitet (B) och originalitet (C). Medel omvandlades till Z-poäng.
Flexibilitetskomponenten
multivariat ANOVA av flexibilitetsåtgärderna uppvisade en allmän effekt för DRD4 7R genotyp F(2,182) = 4.38, p < 0,05. Signifikanta univariata huvudeffekter för DRD4 genotyp erhölls för aut flexibilitet F (1,183) = 4.57, p < 0.05 och för Torrance flexibilitet F(1,183) = 6.95, p < 0.01. Som visas i Figur 1b uppvisade individer i 7R-allelgruppen i båda fallen lägre flexibilitet jämfört med individer i no7-gruppen (Tabell 2).
Originalitetskomponenten
multivariat ANOVA av originalitetsåtgärderna uppvisade inga signifikanseffekter, F(2,182) = 2,06, p = 0.13, Även om poängen indikerade lägre originalitet för 7R-allelen i DRD4 (se figur 1C, Tabell 2 för detaljer).
könsskillnader
diskussion
dt-uppgifter kräver att deltagarna tillhandahåller flera lösningar på ett givet problem. I de uppgifter som användes här var deltagarna tvungna att tänka på många alternativa möjliga användningsområden för vardagliga föremål (i AUT) och att rita flera ritningar som innehåller en specifik form (cirkel) (i Torrance-uppgiften). Resultaten som presenteras här visar att individer med DRD4 7R-allelen uppvisar lägre kreativitet mätt med DT. Med tanke på att 7R-allelen har associerats med dysfunktionell impulsivitet och dålig hämning (Congdon et al., 2008; Dreber et al., 2009), det faktum att individer med 7R-allelen uppvisar lägre kreativitet kan förklaras av deras oförmåga att undertrycka eller hämma uppenbara (vanliga) svar, vilket indikeras av lägre originalitetspoäng. Lägre flexibilitet och flytpoäng kan indikera att när ett gemensamt svar har givits blir det svårare att byta till en ny kategori (vilket indikeras av låga flexibilitetspoäng), vilket leder till Färre svar (vilket indikeras av låg flytpoäng). Ytterligare bevis för förhållandet mellan kreativitet och impulsivitet kan ses i föreningen av 7R-allelen med ADHD (Li et al., 2006), liksom med ökad impulsivitet hos ADHD-patienter (Langley et al., 2004). Intressant nog har kreativitet kopplats till DA-systemet vid schizofreni (Eysenck, 1993). Även om flera studier visar att patienter med schizofreni och schizotypisk personlighet uppvisar hög kreativitet (O ’ Reilly et al., 2001; Folley och Park, 2005), nya modeller av schizofreni och kreativitet stöder en inverterad U-formad förening där kreativiteten kan vara högre med lågmåttig schizotypisk egenskap, men minskar när svårighetsgraden av psykopatologi i schizofreni ökar (Tsakanikos och Claridge, 2005; Stoneham och Coughtrey, 2009; Nelson och Rawlings, 2010). Verkligen, i en nyligen genomförd studie Jaracz et al. (2012) rapporterade att patienter med schizofreni visar minskad kreativitet jämfört med kontroller. Vidare rapporterade författarna att nedsatt kreativitet var förknippad med låg kognitiv flexibilitet och impulsivitet, vilket återspeglas av deras låga poäng på Wisconsin Card Sorting Test (WCST). Dessa resultat är i linje med tidigare forskning som visar att patienter med kronisk schizofreni uppvisar lägre kreativitet, vilket i fallet med kreativ flyt medierades av poäng på verkställande kontrolluppgifter och i fallet med originalitet inte var (Abraham et al., 2007). Sammantaget, medan mild funktionell impulsivitet kan förbättra kreativiteten bland patienter med schizotypiska egenskaper, särskilt när det gäller att generera ideer som skiljer sig från de givna exemplen (Abraham och Windmann, 2008), kan den dysfunktionella impulsiviteten som observeras vid svår schizofreni dämpa kreativa förmågor.
resultaten av den aktuella studien stöder förhållandet mellan kreativitet och varianter av Da-genetiska polymorfier (Reuter et al., 2006; Runco et al., 2011). Dessutom visar resultaten i vårt prov att den huvudsakliga effekten av DRD4-7R-allelen på kreativitet kan bero på dess inverkan på flexibilitet. Flera studier har kopplat kognitiv flexibilitet med DA-vägar (Ashby och isen, 1999; Dreisbach och Goschke, 2004; m Obbller et al., 2007). Det har postulerats att aktivering av D2-liknande receptorer (d.v. s. D2, D3 och D4) minskar Gabaergisk hämning och underlättar aktivering av flera representationer, vilket ökar flexibiliteten som manifesteras av uppsättningsförskjutningsförmåga (M., 2007; Seamans och Robbins, 2010). Dessutom har D2-liknande receptorer kopplats till flexibel integration av ny information (Durstewitz och Seamans, 2008). I linje med detta, Reuter et al. (2006) fann en koppling mellan dopaminreceptorn D2 och kreativitet mätt med ett sammansatt index, men huruvida flexibilitet spelade en viktig roll i dessa resultat kan inte bestämmas. Dessutom, Runco et al. (2011) rapporterade skillnader i kreativ flyt bland bärare av flera dopamingener, bland dem DRD4. Det framgår emellertid inte av deras rapport vilken grupp som uppvisar större kreativa förmågor. Det är viktigt att notera att i motsats till de resultat som presenteras här, Runco et al. (2011) hittade ingen koppling mellan mått på flexibilitet mätt med DT-uppgifter och DRD4. Denna inkonsekvens kan förklaras av skillnader i populationer som används och gruppstorlekar eller av skillnader i allelfrekvenser inom den samplade populationen. Dessutom finns det bevis som tyder på att sambandet mellan dopamin och DT kanske inte är linjärt (Akbari Chermahini och Hommel, 2010, 2012), vilket indikerar att en interaktion mellan flera DA-gener kan påverka de enskilda resultaten av varje genetisk variation i samband med DT. Dessutom har flera studier pekat på det faktum att da-vägar kan vara könskänsliga (Tammim Exceptioni och M. A. A. A., 2011; Wang et al., 2012). Zhong et al. (2010) fann till exempel att kön modulerade sambandet mellan DRD4 exon3 polymorfism och känsla av rättvisa. I den aktuella studien hittade vi inte en interaktionseffekt mellan DRD4-7R och kön, vilket indikerar att kön inte har en modulerande effekt på sambandet mellan DRD4 och kreativitet i vårt urval. Dessutom visar resultaten som presenteras här en brist på könsskillnader i kreativitet, och i synnerhet i DT (för granskning, se Baer och Kaufman, 2008).
Sammanfattningsvis föreslår vi i den aktuella studien att sambandet mellan DRD4 och kreativitet huvudsakligen påverkas av flexibilitet. Resultaten av den aktuella studien är i linje med tidigare resultat som pekar på Da-systemets engagemang i kreativitet och lägger till den ackumulerande kunskapskroppen som tyder på genetiska influenser i kreativitet.
det finns flera begränsningar i studien som bör erkännas. Först undersökte den aktuella studien endast sambandet mellan en genetisk variation i DA—systemet—DRD4-och DT. Det finns emellertid andra viktiga polymorfier i striatal DA-systemet som kan associeras med DT och kan bidra både på ett fristående sätt såväl som i gen-gen interaktioner. En sådan polymorfism är dopamintransportören (DAT1), som har visat sig vara associerad med hämning av retur (Colzato et al., 2010) samt med impulsivitetsåtgärder (Gizer och Waldman, 2012). Vidare, som nämnts ovan, är en annan möjlighet som inte undersöktes här de möjliga gen-geninteraktionseffekterna på DT. Akbari Chermahini och Hommel (2010) rapporterade en olinjär koppling mellan en markör för striatal dopamin och DT, vilket indikerar att förhållandet kan vara mer komplext, så att en polymorfism av en gen kan påverka DT genom medling av en annan genpolymorfism. Således bör framtida studier överväga att undersöka interaktionen mellan dessa två polymorfier i samband med flexibilitet och DT.
intressekonflikt uttalande
författarna förklarar att forskningen genomfördes i avsaknad av kommersiella eller finansiella relationer som kan tolkas som en potentiell intressekonflikt.
Akbari Chermahini, S. och Hommel, B. (2010). (B) länken mellan kreativitet och dopamin: spontana ögonblinkhastigheter förutsäger och dissocierar divergerande och konvergerande tänkande. Kognition 115, 458-465. doi: 10.1016/j.kognition.2010.03.007
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
kyler, R. (2008). Roll dopamin i motiverande och kognitiv kontroll av beteende. Neuroscientist 14, 381-395. doi: 10.1177 / 1073858408317009
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Frank, M. J. (2005). Dynamisk dopaminmodulering i basala ganglier: en neurocomputational redogörelse för kognitiva underskott i medicinerad och icke-medicinsk Parkinsonism. J. Cogn. Neurovetenskap. 17, 51–72. doi: 10.1162 / 0898929052880093
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Healey, D. och Rucklidge, JJ (2006). En undersökning av förhållandet mellan ADHD-symptomatologi, kreativitet och neuropsykologisk funktion hos barn. Barn Neuropsychol. 12, 421–438. doi: 10.1080 / 09297040600806086
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Runco, Ma och Richards, R. (1997). Eminent kreativitet, vardaglig kreativitet och hälsa. Greenwich, CT: Ablex.
Torrance, E. (1974). Skillnader är inte underskott. Lära. Coll. Rec. 75, 471–487.