- värdspecifik interaktion mellan Rhizobium och Växtpartners
- bestämd och obestämd Nodulutveckling
- tillväxt av symbiotiska celler involverar förstärkning av värdgenomet genom Endoredupliceringscykler
- olika öden för kvävefixerande bakterier
- Värdpeptider styr Bakteroiddifferentiering
- NCR247: ett exempel för multi-Target Host Effector
- slutsats
- intressekonflikt uttalande
- erkännanden
värdspecifik interaktion mellan Rhizobium och Växtpartners
bakterierna som bildar kvävefixerande symbios med baljväxtväxter som tillhör olika grupper av sackaros-och tuberkulosproteobakterier kallas kollektivt rhizobia (Chen et al., 2003; MacLean et al., 2007). Många av de Proteobakterier är engagerade i långsiktiga interaktioner med högre eukaryoter. Dessa interaktioner sträcker sig från ytkolonisering genom fakultativa symbiotiska förhållanden för att förplikta intracellulär patogen eller endosymbiont livsstil. De symbiotiska gener som krävs för nodulbildning, värdcellinfektion och kvävefixering har förvärvats genom lateral genöverföring som är den primära källan till genetisk mångfald av rhizobia. Därför kan rhizobia vara närmare besläktad med patogener (såsom Agrobacterium eller Brucella) än till varandra. Rhizobia tenderar att ha stora genom (upp till 10.5 Mbp) som i snabbväxande rhizobia sprids på flera replikoner (MacLean et al., 2007). Till exempel har Sinorhizobium meliloti, endosymbionten av Medicago-arter, ett tri-delat genom; en 3,65 Mbp-kromosom och två megaplasmider, pSymA och pSymB (1,35 och 1,68 Mbp) som båda är oumbärliga och bär majoriteten av symbiotiska gener. Emellertid innehåller många S. meliloti-stammar ytterligare hjälpmedelstora plasmider och sålunda kan S. meliloti-genomet innehålla upp till 9000 gener (Barnett et al., 2001; Capela et al., 2001). I motsats till rhizobia har obligatoriska endosymbionter av insekter vanligtvis ett starkt reducerat (160-450 Kbp) genom som säkerställer deras multiplikation och koder för några specifika biosyntetiska vägar inklusive de som uppfyller värdens behov (Moran et al., 2008; Price et al., 2011). Dessa otroligt reducerade genom förstärks ändå och kompenserar det minskade genomet med ett polyploid DNA-innehåll.
rhizobias växtpartners tillhör familjen Leguminosae / Fabaceae. Kvävefixerande symbios har utvecklats i flera linjer, men inte alla baljväxter bildar symbios. Hittills är 12 000 nodulerade baljväxtarter kända och var och en har sin egen Rhizobiumpartner(er). Symbiosen utlöses av kvävehushållning av värdväxten som måste välja sin Rhizobiumpartner från miljarder bakterier i rhizosfären. Detta uppnås genom utsöndring av flavonoidsignalmolekyler från roten som fungerar som kemoattraktiva men viktigast som inducerare av Rhizobium noduleringsgenerna (Oldroyd, 2013). Dessa gener krävs för produktion av bakteriella signalmolekyler; Nodfaktorerna (NFs) som utlöser nodulutvecklingsprogrammet i värdväxten (Walker och Downie, 2000). NFs är lipochitooligosackaridmolekyler som bär värdspecifika substitutioner på de terminala sockerresterna och karakteristiska lipidkedjor, som känns igen av Värdreceptorer av LysM-typ och krävs både för nodulutveckling och bakteriell infektion. Intressant är att den antika symbiosen av landväxter med arbuscular mycorrhizal (AM) svampar fungerar med liknande lipochitooligosackaridsignalmolekyler, Myc-faktorerna som uppfattas av liknande men olika lysm-typreceptorer som NFs (Abdel-Lateif et al., 2012; Oldroyd, 2013). Myc-faktorerna och NFs aktiverar en gemensam signalväg men efter involveringen av de vanliga symbiotiska generna som konserveras i växter avviker vägarna; en som leder till nodulering, den andra för AM-symbios.
Växtinfektion och nodulbildning är invecklade processer; Nodfaktorer spelar olika roller i nodulorganogenes och rothårinfektion. Dessutom, förutom Nodfaktorer, är olika bakteriella ytpolysackarider avgörande för effektiv infektion (Fraysse et al., 2003). I de flesta baljväxter kommer rhizobia in i värden via rothåren där genom invagination av plasmamembranet bildas en infektionstråd (IT) som innehåller de multiplicerande bakterierna och växer mot rotbarken. Ett mindre frekvent och gammalt infektionssätt sker via sprickor på rotytan hos vissa baljväxter.
bestämd och obestämd Nodulutveckling
Nodulutveckling kräver mitotisk reaktivering av kortikala celler som leder till nodulär primordiumbildning som sedan differentierar till kvävefixerande rotknutor som ger mikroaerobiskt tillstånd i den centrala zonen för funktion av det syrekänsliga kväveasenzymet i bakterioiderna. Beroende på den övergående och ihållande naturen hos värdcellsproliferation kan nodulerna vara antingen bestämda eller obestämda typ (Terpolilli et al., 2012; Kondorosi et al., 2013). Bestämda knölar har ingen meristem och innehåller homogen population av symbiotiska celler. Bestämda knölar utvecklas till exempel på Phaseolus vulgaris och Lotus japonicus rötter.
tvärtom upprätthålls den aktiva celldelningen i de obestämda nodulerna. En nodulär meristem är närvarande i den apikala regionen (zon I) som genom konstant generering av nya celler framkallar kontinuerlig tillväxt och långsträckt nodulär form. Cellerna som lämnar meristemet delar sig inte längre och går in i en differentieringsfas. Infektionstråden frigör bakterierna i de submeristematiska cellerna, som differentierar gradvis längs 12-15-cellskikten i infektionszonen (zon II), vilket leder till utveckling av kvävefixerande symbiotiska celler i nodulzon III (Figur 1; Franssen et al., 1992). Medicago sativa, M. truncatula, Vicia sativa och Pisum sativum är exempel på växter som bildar obestämda knölar.
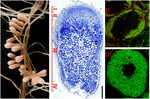
figur 1. Struktur av kvävefixerande rotknutor bildade i S. meliloti – M. truncatula symbios. De olika nodulzonerna anges på den längsgående nodulsektionen: (i) meristem, (II) infektionszon, (III) kvävefixeringszon, (IV) senescenszon. Symbiotiska celler i zon II innehåller de differentierande endosymbionerna medan värdcytoplasman i zon III är fullpackad med långa kvävefixerande bakterier. Endosymbionter färgade med Syto9 har grön fluorescens.
tillväxt av symbiotiska celler involverar förstärkning av värdgenomet genom Endoredupliceringscykler
Extrem växtcellförstoring kan observeras i både de bestämda och obestämda nodulerna. Cytoplasman hos en kvävefixerande symbiotisk cell är värd för cirka 50 000 bakterier. För att rymma ett så stort antal endosymbionter växer värdcellerna. I M. truncatula noduler är volymen av kvävefixerande celler 80 gånger större än den för de diploida meristematiska cellerna. Tillväxten av infekterade celler sker stegvis i zon II och är konsekvensen av upprepad endoreduplicering (ER) av genomet utan mitos. I zon II är cellcykelmaskineriet fortfarande aktivt men bristen på mitotiska cykliner hämmar mitos och omvandlar de mitotiska cyklerna till endoredupliceringscykler (Cebolla et al., 1999). Detta uppnås genom cellcykelomkopplaren CCS52A-protein som genom förstörelsen av de mitotiska cyklinerna inducerar upprepade omgångar av genomduplikation som leder till bildandet av gradvis växande polyploida celler (Roudier et al., 2003; Kondorosi och Kondorosi, 2004). I Medicago-arter kan ploidi-nivåerna nå 64C som representerar 64-faldigt högre DNA-innehåll jämfört med haploida celler (C motsvarar haploid DNA-innehåll; Vinardell et al., 2003). Nedreglering av CCS52A i M. truncatula hade ingen effekt på primordiumbildning men var skadlig för noduldifferentiering vilket indikerar att er-cyklerna och bildandet av stora högpolyploida celler är väsentliga för nodulfunktion (Vinardell et al., 2003). Intressant är att kortikala celler som innehåller am-svampar också är polyploida, liksom de nematodmatande jätterotcellerna (Favery et al., 2002; Genre et al., 2008). På liknande sätt är insektssymbiotiska celler, bakteriocyterna som har intracellulära endosymbionter också stora och polyploida (Nakabachi et al., 2010). I angiospermväxter är polyploidi frekvent och det specifika ärvda mönstret av polyploidi i olika organ, vävnader och celltyper tyder på att det kan vara en viktig källa till värdcellernas specialiserade fysiologi (Nagl, 1976; Edgar et al., 2014). Förutom celltillväxt kan de multipla genkopiorna, brist på kromosomkondensation bidra till högre transkriptionella och metaboliska aktiviteter. Associering av polyploidi med olika cellfunktioner antyder emellertid en inverkan av polyploidi också på nukleosomernas arkitektur och på epigenomet som styr aktivering eller förtryck av specifika genomiska regioner. Följaktligen verkar det polyploida genominnehållet i symbiotiska celler vara en förutsättning för noduldifferentiering och för uttryck av de flesta symbiotiska värdgener (Maunoury et al., 2010).
olika öden för kvävefixerande bakterier
bakterierna som frigörs från IT finns i värdcytoplasman som organellliknande strukturer, kallade symbiosomer. Bakterierna har ingen direkt kontakt med cytoplasma eftersom de omges av ett peribacteroidmembran, även känt som symbiosommembran (SM). Bakterioiden, SM och utrymmet mellan dem innefattar symbiosomen (Catalano et al., 2004). SM under dess bildning återspeglar dess plasmamembran ursprung, senare modifieringar av dess sammansättning öppnar nya, specialiserade roller vid värd-endosymbiont-gränssnittet (Limpens et al., 2009; Ivanov et al., 2012; Brear et al., 2013; Sinharoy et al., 2013). Bakterioiderna multiplicerar i de växande värdnodulcellerna till en viss celldensitet, anpassar sig till endosymbiotiska livsstilar och mikroaerobiska förhållanden och mognar till kvävefixerande bakterier. Bakteriernas form och fysiologi kan dock vara påfallande olika i de olika baljväxterna. I vissa baljväxtvärdar har kvävefixerande bakterier samma morfologi som odlade celler; denna typ av bakterier kan återgå till den fritt levande formen. I andra föreningar omvandlas bakterioiderna irreversibelt till polyploida, förstorade, icke-odlingsbara endosymbionter. Dessa terminalt differentierade bakterier kan vara långsträckta och till och med grenade och 5 – till 10-faldigt längre än de fritt levande cellerna eller kan vara sfäriska från 8 till minst 20-faldigt förstärkt genom beroende på värden (Mergaert et al., 2006; Nakabachi et al., 2010). Terminal differentiering av bakterier är värdstyrd, utvecklad i flera grenar av Leguminosae-familjen som indikerar värdfördel och sannolikt högre symbiotisk prestanda (Oono et al., 2010). Terminal bacteroiddifferentiering är den bästa belysta i S. meliloti – M. truncatula symbios. I M. truncatula noduler, de mest synliga händelserna av terminal bacteroiddifferentiering förekommer i zon II. multiplikation av bacteroids stannar i mitten av zon II där cellförlängning och enhetlig förstärkning av flera replikoner genom endoredupliceringscykler börjar. Längs 2-3 cellskikt vid gränsen till zon II och III (kallad interzon) plötslig tillväxt av bakterier är synlig och når praktiskt taget sin slutliga storlek, men kvävefixering sker endast i zon III.
Värdpeptider styr Bakteroiddifferentiering
jämförelse av nodultranskriptomer av baljväxter med reversibel och irreversibel bakteroiddifferentiering avslöjade förekomsten av flera hundra små gener som endast var närvarande i genomet hos de värdväxter där bakteroiddifferentiering var terminal. I M. truncatula producerar nodulcellerna minst 600 nodulspecifika symbiotiska peptider (symPEPs). SymPEP-generna aktiveras endast i S. meliloti-infekterade polyploida symbiotiska celler (Kevei et al., 2002; Mergaert et al., 2003), men vissa uppsättningar vid det tidigare, andra under de senare stadierna av nodulutveckling. En stor del, mer än 500 gener kodar nodulspecifika cysteinrika (NCR) peptider (Mergaert et al., 2003; Alunni et al., 2007; Nallu et al., 2014). NCR-peptiderna är riktade mot bakteroiderna och när deras leverans till endosymbionterna blockerades avskaffades bakteroiddifferentiering vilket visar att peptiderna är ansvariga för terminal differentiering av S. meliloti-bakterier (Van de Velde et al., 2010). Den höga sekvensvariationen och de karakteristiska uttrycksmönstren för NCR-gener föreslår mångfald i deras funktioner, verkningssätt och bakteriemål vid olika stadier av bakteroidmognad (Figur 2). Men varför producerar värdcellen en arsenal av NCRs? Vad kan vara fördelen med en så mångsidig peptidrepertoar? Är det nödvändigt för värdens interaktion med olika bakterier? De symbiotiska partnerna till M. truncatula är S. meliloti och S. medicae, men i jorden finns det otaliga stamvarianter av båda arterna. M. truncatula representeras också av många olika ekotyper och anslutningar som skiljer sig åt i antal, sekvenser och uttrycksprofil för NCR-gener och i deras symbiotiska interaktioner med olika S. meliloti-och S. medicae-stammar (Nallu et al., 2014; Roux et al., 2014). Medan en nodul innehåller en enda bakterietyp, kan de olika nodulerna på samma rotsystem ha distinkta bakteriepopulationer. Det är möjligt att växten som känner igen de olika endosymbionerna manipulerar dem med en stamspecifik repertoar av peptider. Dessa skillnader kan lägga till en ytterligare kontrollnivå för värd-symbiont-specificitet och därmed för noduleringseffektivitet.

figur 2. Differentiellt uttryck av sympepgener i M. truncatula noduler. Svart signal: in situ hybridisering, blå signal: Gus aktivitet av symPEP promotor-GUS fusioner i transgena knölar.
även om symPEPs representerar unika peptidklasser, liknar deras strukturer antimikrobiella peptider (ampere). Ampere med brett spektrum av mikrobiell celldödande aktivitet är oftast katjonisk provokerande celldöd genom porbildning, membranstörning och därmed lys av mikrobiella celler. Det faktum att celldelningsförmågan definitivt förloras under endosymbiontdifferentiering indikerar att åtminstone vissa symPEPs har antimikrobiella aktiviteter. Behandling av bakterier med syntetiska katjoniska NCRs provocerade faktiskt snabb och effektiv dosberoende eliminering av olika gramnegativa och grampositiva bakterier inklusive viktiga humana och växtpatogener (Van de Velde et al., 2010; Tiricz et al., 2013). Denna ex-planta-dödande effekt korrelerad med permeabilisering av mikrobiella membran, men symPEPs i deras naturliga miljö – i nodulcellerna – genomtränger inte bakteriemembranen och dödar inte endosymbionerna. Mest sannolikt är peptidkoncentrationerna i nodulerna signifikant lägre än de som appliceras i in vitro-analyserna. Dessutom produceras katjoniska peptider tillsammans med anjoniska och neutrala peptider i samma cell, och möjlig kombination av några tiotals eller hundratals peptider med olika laddning och hydrofobicitet kan neutralisera den direkta bakteriedödande effekten av de katjoniska peptiderna.
involveringen av ampere eller AMP-liknande peptider är inte unik för Rhizobium-legume symbios. I weevil Sitophilus producerar de symbiotiska cellerna den antimikrobiella peptiden coleoptericin-A (ColA) som framkallar utvecklingen av jätte filamentösa endosymbionter genom att hämma celldelning och skyddar de närliggande insektsvävnaderna från bakteriell invasion (Login et al., 2011). I detta system är en enda peptid tillräcklig för differentiering av den obligatoriska vertikalt överförda endosymbiont till skillnad från noduler som arbetar med hundratals symPEPs och kan vara värd för otaliga stamvarianter som deras endosymbionter. I aphid-Buchnera-symbiosen producerar värdcellerna också bakteriocytspecifika peptider inklusive cysteinrika peptider (BCR) som liknar Medicago NCR-peptiderna, men funktionerna hos dessa symbiotiska peptider har inte rapporterats ännu (Shigenobu och Stern, 2013).
NCR247: ett exempel för multi-Target Host Effector
transkriptomanalys av M. truncatula noduler i olika stadier av deras utveckling, lasermikrodissektion av nodulregioner, in situ hybridisering, immunolokalisering av utvalda peptider och symPEP promotor-reporter-genfusioner i transgena noduler möjliggör kartläggning av verkan av enskilda peptider i symbiotiska celler från tidig infektion till sent kvävefixeringstillstånd. NCR247 uttrycks i de äldre cellskikten i zon II och i interzonen där bakteriell celldelning stannar och anmärkningsvärd förlängning av endosymbionterna uppträder (Farkas et al., 2014). Denna lilla katjoniska peptid dödade effektivt olika mikrober in vitro och in silico-analysen indikerade dess extrema proteinbindningsförmåga. FITC-märkt NCR247 gick in i bakteriecytosolen där dess interaktioner med många bakterieproteiner var möjliga. Bindningspartners identifierades genom behandling av S. meliloti-bakterier eller bakterier med StrepII/FLAGGMÄRKTA peptider följt av affinitetskromatografi och identifiering av interagerande partners med LC-MS/MS och västerländsk analys (Farkas et al., 2014).
en av interaktorerna var FtsZ – celldelningsproteinet som spelade en avgörande primär roll i celldelning. Ett antal antibiotiska peptider är kända för att utöva bakteriedödande eller bakteriostatisk effekt genom interaktionen med FtsZ, vilket hämmar dess polymerisation och därigenom hindrar korrekt Z-ring och septumbildning (Handler et al., 2008). NCR247 samrenades med FtsZ från bakteriell cytoplasma och visade sig störa septumbildning. NCR035 uppvisar in vitro också bakteriedödande effekt och produceras i samma symbiotiska celler som NCR247 ackumuleras vid division septum som indikerar samtidig eller på varandra följande verkan av dessa peptider och utveckling av flera värdstrategier för att hämma endosymbiontproliferation. En annan studie visade att uttryck av viktiga celldelningsgener, inklusive gener som krävs för Z-ringfunktion, dämpades starkt i celler behandlade av NCR247 (Penterman et al., 2014). Förbehandling av bakterier med subletala ncr247-koncentrationer avskaffade lokalisering av FITC-NCR035 till septum och provocerad cellförlängning (Farkas et al., 2014).
ribosomala proteiner var de vanligaste ncr247-interagerande partnerna. NCR247 observerades starkt hämma bakteriell proteinsyntes på ett dosberoende sätt både in vivo och in vitro (Farkas et al., 2014). Dessa resultat antydde att ett läge för ncr247-peptidverkan binder till ribosomerna både i bakterieceller och bakteroider. Intressant nog observerades ett förändrat mönster och minskad komplexitet hos de interagerande proteinerna i bakterioiderna. Följaktligen var den allmänna expressionsnivån för ribosomala proteiner i genomsnitt 20 gånger lägre i bakterioiderna än i de fritt levande cellerna med olika relativa överflöd av transkript av enskilda ribosomala proteiner. Ribosomdiversifiering i bakterier kan ha en betydande roll genom att bidra till den avancerade översättningen av specifika proteiner och därigenom stödja den specialiserade, energikrävande fysiologin av mycket riklig kvävefixeringsfunktion.
GroEL chaperon var också en direkt samverkande partner till NCR247 (Farkas et al., 2014). Av de 5 GroEL-proteinerna är GroEL1 eller GroEL2 tillräcklig för överlevnad medan GroEL1 uttryckt på hög nivå i nodulen är avgörande för symbios (Bittner et al., 2007). Det behövs för fullständig aktivering av noduleringsgenerna och montering av kväveas-komplexet. GroEL har extrem funktionell mångsidighet genom att interagera med hundratals proteiner. Ncr247-GroEL1-interaktionen kan få konsekvenser direkt på GroEL men indirekt också på Groelsubstraten och tillhörande biologiska processer. Frånvaro av GroEL1 allvarligt påverkad bakteriell infektion och underhåll och differentiering av bakterier som visar ett allmänt behov av GroEL1 i alla stadier av kvävefixeringsnodulutveckling.
involveringen av GroEL-och värdpeptider i mikrobe-värdinteraktioner är inte unik för Rhizobium-legume symbios. I weevil samverkar symbiotiska celler coleoptericin-a (ColA) också med GroEL (Login et al., 2011). GroEL spelar också en viktig roll i underhållet av endosymbionter (Moran, 1996; Kupper et al., 2014). Eftersom de flesta symbiotiska system ännu är outforskade och genomiska och proteomiska verktyg med hög genomströmning är bara nyligen tillgängliga, kan vi bara förutsäga att värdpeptider-medierad endosymbiontdifferentiering, likaså genomförstärkning av värdceller och terminalt differentierade endosymbionter är allmänna strategier för symbios.
slutsats
symbiotiska och patogena bakterier använder liknande metoder för att interagera med sina värdar och att överleva inom värdceller, även om resultaten av dessa interaktioner är påfallande olika. Växter och djur kan generera medfödda immunsvar mot mikroorganismer vid uppfattningen av MAMPs (mikroorganismassocierade molekylära mönster). Denna uppfattning resulterar i aktivering av signalkaskader och produktion av antimikrobiella effektorer. AMP-liknande värdpeptider såsom M. truncatula NCR-peptider eller Weevil ColA antimikrobiell peptid spelar viktiga och mångfacetterade roller för att kontrollera multiplikationen och differentieringen av endosymbionter, vilket begränsar närvaron av bakterier till de symbiotiska cellerna. Således använder värdorganismer dessa effektorpeptider för att tämja och till och med anställa utvalda mikrobiella invaderare för service.
intressekonflikt uttalande
författarna förklarar att forskningen genomfördes i avsaknad av kommersiella eller finansiella relationer som kan tolkas som en potentiell intressekonflikt.
erkännanden
arbetet i våra laboratorier stöds av Europeiska forskningsrådets” SYM-BIOTICS ” avancerade bidrag till Chubbiva Kondorosi (bidragsnummer 269067) och av T USBI-4.2.2.A-11/1 / KONV-2012-0035 stöds av Europeiska unionen och samfinansieras av Europeiska socialfonden.
Abdel-Lateif, K., Bogusz, D. och Hocher, V. (2012). Rollen av flavonoider i etableringen av växtrötter endosymbioser med arbuscular mycorrhiza svampar, rhizobia och Frankia bakterier. Växt Signal. Uppför dig. 7, 636–641. doi: 10.4161 / psb.20039
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Alunni, B., Kevei, Z., Redondo-Nieto, M., Kondorosi, A., Mergaert, P., and Kondorosi, E. (2007). Genomic organization and evolutionary insights on GRP and NCR genes, two large nodule-specific gene families in Medicago truncatula. Mol. Plant Microbe Interact. 20, 1138–1148. doi: 10.1094/MPMI-20-9-1138
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Barnett, M. J., Fisher, R. F., Jones, T., Komp, C., Abola, A. P., Barloy-Hubler,F., et al. (2001). Nukleotidsekvens och förutsagda funktioner för hela Sinorhizobium meliloti pSymA megaplasmid. Proc. Natl. Acad. Sci. USA 98, 9883-9888. doi: 10.1073/pnas.161294798
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Bittner, A. N., Foltz, A. och Oke, V. (2007). Endast en av fem groelgener krävs för livskraft och framgångsrik symbios i Sinorhizobium meliloti. J. Bakteriol. 189, 1884–1889. doi: 10.1128 / JB.01542-06
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Brear, E. M., Day, D. A., and Smith, P. M. (2013). Iron: an essential micronutrient for the legume-Rhizobium symbiosis. Front. Plant Sci. 4:359. doi: 10.3389/fpls.2013.00359
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Catalano, C. M., Lane, W. S., and Sherrier, D. J. (2004). Biochemical characterization of symbiosome membrane proteins from Medicago truncatula root nodules. Electrophoresis 25, 519–531. doi: 10.1002/elps.200305711
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Capela, D., Barloy-Hubler, F., Gouzy, J., Bothe, G., Ampe, F., Batut, J.,et al. (2001). Analys av kromosomsekvensen av baljväxter symbiont Sinorhizobium meliloti stam 1021. Proc. Natl. Acad. Sci. USA 98, 9877-9882. doi: 10.1073/pnas.161294398
Pubmed Abstract / Pubmed fulltext / CrossRef fulltext
Cebolla, A., Vinardell, J. M., Kiss, E., ol ol, B., Roudier, F., Kondorosi, A.,et al. (1999). Den mitotiska hämmaren ccs52 krävs för endoreduplicering och ploidiberoende cellförstoring i växter. EMBO J. 18, 4476-4484. doi: 10.1093 / emboj / 18.16.4476
CrossRef fulltext
Chen, W. M., Moulin, L., Bontemps, C., Vandamme, P., B Excepilna, G. och Boivin-Masson, C. (2003). Baljväxtsymbiotisk kvävefixering med hjälp av OC-Proteobakterier är utbredd i naturen. J. Bakteriol. 185, 7266–7272. doi: 10.1128 / JB.185.24.7266-7272.2003
CrossRef fulltext
Edgar, ba, Zielke, N. och Gutierrez, C. (2014). Endocykler: en återkommande evolutionär innovation för post-mitotisk celltillväxt. Nat. Pastor Mol. Cell. Biol. 15, 197–210. doi: 10.1038/nrm3756
Pubmed Abstrakt | Pubmed Full Text | CrossRef Full Text
Farkas, A., Maróti, G., Dürgõ, H., Györgypál, Z., Lima, R. M., Medzihradszky, K. F.,et al. (2014). Medicago truncatula symbiotisk peptid NCR247 bidrar till bakteroiddifferentiering genom flera mekanismer. Proc. Natl. Acad. Sci. USA 111, 5183-5188. doi: 10.1073/pnas.1404169111
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Favery, B., Complainville, A., Vinardell, J. M., Lecomte, P., Vaubert, D., Mergaert, P.,et al. (2002). The endosymbiosis-induced genes ENOD40 and CCS52a are involved in endoparasitic-nematode interactions in Medicago truncatula. Mol. Plant-Microbe Interact. 15, 1008–1013. doi: 10.1094/MPMI.2002.15.10.1008
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Franssen, H. J., Vijn, I., Yang, W. C., and Bisseling, T. (1992). Developmental aspects of the Rhizobium-legume symbiosis. Plant Mol. Biol. 19, 89–107. doi: 10.1007
CrossRef Full Text
Fraysse, N., Couderc, F., and Poinsot, V. (2003). Surface polysaccharide involvement in establishing the Rhizobium–legume symbiosis. Eur. J. Biochem. 270, 1365–1380. doi: 10.1046/j.1432-1033.2003.03492.x
CrossRef Full Text
Genre, A., Chabaud, M., Faccio, A., Barker, D. G., and Bonfante, P. (2008). Prepenetrationsapparatmontering föregår och förutsäger koloniseringsmönstren för arbuscular mycorrhizal svampar inom rotbarken hos både Medicago truncatula och Daucus carota. Växtcell 20, 1407-1420. doi: 10.1105/tpc.108.059014
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Handler, AA, Lim, Je och Losick, R. (2008). Peptidhämmare av cytokinesis under sporulering i Bacillus subtilis. Mol. Mikrobiol. 68, 588–599. doi: 10.1111/j.1365-2958.2008. 06173.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Ivanov, S., Fedorova, E. E., Limpens, E., De Mita, S., Genre, A., Bonfante, P.,et al. (2012). Rhizobium–legume symbiosis shares an exocytotic pathway required for arbuscule formation. Proc. Natl. Acad. Sci. U.S.A. 109, 8316–8321. doi: 10.1073/pnas.1200407109
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Kevei, Z., Vinardell, J. M., Kiss, G. B., Kondorosi, A., and Kondorosi, E. (2002). Glycinrika proteiner kodade av en nodulspecifik genfamilj är inblandade i olika stadier av symbiotisk nodulutveckling i Medicago spp. Mol. Växtmikroben Interagerar. 15, 922–931. doi: 10.1094/MPMI.2002.15.9.922
Pubmed Abstrakt | Pubmed fulltext/CrossRef fulltext
Kondorosi, E. och Kondorosi, A. (2004). Endoreduplicering och aktivering av det anafasfrämjande komplexet under symbiotisk cellutveckling. FEBS Lett. 567, 152–157. doi: 10.1016/j.febslet.2004.04.075
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Kondorosi, E., Mergaert, P. och Kereszt, A. (2013). Ett paradigm för endosymbiotiskt liv: celldifferentiering av Rhizobiumbakterier provocerade av värdväxtfaktorer. Annu. Rev. Mikrobiol. 67, 611–628. doi: 10.1146 / annurev-micro-092412-155630
CrossRef fulltext
Kupper, M., Gupta, Sk, Feldhaar, H. och Gross, R. (2014). Mångsidiga roller hos chaperonin GroEL i interaktioner mellan mikroorganismer och insekter. FEMS Microbiol. Lett. 353, 1–10. doi: 10.1111 / 1574-6968.12390
Pubmed Abstrakt | Pubmed fulltext | CrossRef fulltext
Limpens, E., Ivanov, S., van Esse, W., Voets, G., Fedorova, E. och Bisseling, T. (2009). Medicago N2-fixerande symbiosomer förvärvar den endocytiska identitetsmarkören Rab7 men försenar förvärvet av vakuolär identitet. Växtcell 21, 2811-2828. doi: 10.1105/tpc.108.064410
Pubmed Abstract / Pubmed fulltext / CrossRef fulltext
Logga in, F. H., Balmand, S., Vallier, A., Vincent-Mon Oubergat, C., Vigneron, A., Weiss-Gayet, M., et al. (2011). Antimikrobiella peptider håller insektsendosymbionter under kontroll. Vetenskap 334, 362-365. doi: 10.1126/vetenskap.1209728
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
MacLean, am, Finan, TM och Sadowsky, MJ (2007). Genomer av de symbiotiska kvävefixerande bakterierna av baljväxter. Växt Physiol. 144, 615–622. doi: 10.1104 / s. 107. 101634
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Maunoury, N., Redondo-Nieto, M., Bourcy, M., Van de Velde, W., Alunni, B., Laporte, P., et al. (2010). Differentiering av symbiotiska celler och endosymbionter är kopplade till två transkriptomomkopplare i Medicago truncatula nodulation. PLoS EN 5: e9519. doi: 10.1371 / tidskrift.pone.0009519
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Mergaert, P., Nikovics, K., Kelemen, Z., Maunoury, N., Vaubert, D., Kondorosi, A.,et al. (2003). En ny familj i Medicago truncatula bestående av mer än 300 nodulspecifika gener som kodar för små, utsöndrade polypeptider med konserverade cysteinmotiv. Växt Physiol. 132, 161–173. doi: 10.1104/pp.102.018192
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Mergaert, P., Uchiumi, T., Alunni, B., Evanno, G., Cheron, A., Catrice, O.,et al. (2006). Eukaryotic control on bacterial cell cycle and differentiation in the Rhizobium-legume symbiosis. Proc. Natl. Acad. Sci. U.S.A. 103, 5230–5235. doi: 10.1073/pnas.0600912103
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Moran, N. A. (1996). Accelererad utveckling och Mullers spärr i endosymbiotiska bakterier. Proc. Natl. Acad. Sci. USA 93, 2873-2878. doi: 10.1073/pnas.93.7.2873
CrossRef fulltext
Moran, na, McCutcheon, jp och Nakabachi, A. (2008). Genomik och utveckling av ärftliga bakteriella symbionter. Annu. Pastor Genet. 42, 165–690. doi: 10.1146/annurev.genet.41.110306.130119
CrossRef fulltext
Nakabachi, A., Koshikawa, S., Miura, T. och Miyagishima, S. (2010). Genomstorlek av Pachypsylla venusta (Hemiptera: Psyllidae) och ploidi av dess bakteriocyt, den symbiotiska värdcellen som har intracellulära mutualistiska bakterier med det minsta cellulära genomet. Tjur. Entomol. Res 100, 27-33. doi: 10.1017 / S0007485309006737
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Nagl, W. (1976). DNA endoreduplication och polyteny förstås som evolutionära strategier. Natur 261, 614-615. doi: 10.1038 / 261614a0
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Nallu, S., Silverstein, K. A., Zhou, P., Young, nd och Vandenbosch, K. A. (2014). Mönster av divergens av en stor familj av nodulcysteinrika peptider i anslutningar av Medicago truncatula. Växt J. 78, 697-705. doi: 10.1111/tpj.12506
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Oldroyd, G. E. (2013). Tala, vän och gå in: signalsystem som främjar fördelaktiga symbiotiska föreningar i växter. Nat. Rev. Mikrobiol. 11, 252–263. doi: 10.1038/nrmicro2990
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Oono, R., Schmitt, I., Sprent, J. I., and Denison, R. F. (2010). Multiple evolutionary origins of legume traits leading to extreme rhizobial differentiation. New Phytol. 187, 508–520. doi: 10.1111/j.1469-8137.2010.03261.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Penterman, J., Abo, R. P., De Nisco, N. J., Arnold, M. F., Longhi, R., Zanda, M.,et al. (2014). Värdväxtpeptider framkallar ett transkriptionssvar för att kontrollera Sinorhizobium meliloti-cellcykeln under symbios. Proc. Natl. Acad. Sci. USA 111, 3561-3566. doi: 10.1073/pnas.1400450111
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
pris, Dr, Duncan, Rp, Shigenobu, S. och Wilson, AC (2011). Genomexpansion och differentiellt uttryck av aminosyratransportörer vid aphid/buchnera symbiotiska gränssnitt. Mol. Biol. Evol. 28, 3113–3126. doi: 10.1093/molbev/msr140
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Roudier, F., Fedorova, E., Lebris, M., Lecomte, P., Györgyey, J., Abad, P.,et al. (2003). The Medicago species A2-type cyclin is auxin regulated and involved in meristem formation but dispensable for endoreduplication-associated developmental programs. Plant Physiol. 131, 1091–1103. doi: 10.1104/pp.102.011122
CrossRef Full Text
Roux, B., Rodde, N., Jardinaud, M. F., Timmers, T., Sauviac, L., Cottret, L.,et al. (2014). En integrerad analys av växt-och bakteriegenuttryck i symbiotiska rotknutor med laserfångande mikrodissektion kopplad till RNA-sekvensering. Växt J. 77, 817-837. doi: 10.1111/tpj.12442
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Shigenobu, S. och Stern, D. L. (2013). Bladlöss utvecklade nya utsöndrade proteiner för symbios med bakteriell endosymbiont. Proc. Biol. Sci. 280, 20121952. doi: 10.1098/rspb.2012.1952
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Sinharoy, S., Torres-Jerez, I., Bandyopadhyay, K., Kereszt, A., Pislariu, C. I., Nakashima, J.,et al. (2013). C2H2-transkriptionsfaktorregulatorn för symbiosomdifferentiering undertrycker transkription av den sekretoriska vägen Gen VAMP721a och främjar symbiosomutveckling i Medicago truncatula. Växtcell 25, 3584-3601. doi: 10.1105/tpc.113.114017
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Terpolilli, J. J., Hood, G. A., and Poole, P. S. (2012). What determines the efficiency of N(2)-fixing Rhizobium-legume symbioses? Adv. Microb. Physiol. 60, 325–389. doi: 10.1016/B978-0-12-398264-3.00005-X
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Tiricz, H., Szûcs, A., Farkas, A., Pap, B., Lima, R. M., Maróti, G.,et al. (2013). Antimicrobial nodule-specific cysteine-rich peptides induce membrane depolarization associated changes in Sinorhizobium meliloti. Appl. Env. Microbiol. 79, 6737–6746. doi: 10.1128/AEM.01791-13
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Van de Velde, W., Zehirov, G., Szatmari, A., Debreczeny, M., Ishihara, H., Kevei, Z.,et al. (2010). Plant peptides govern terminal differentiation of bacteria in symbiosis. Science 327, 1122–1126. doi: 10.1126/science.1184057
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Vinardell, J. M., Fedorova, E., Cebolla, A., Kevei, Z., Horvath, G., Kelemen, Z.,et al. (2003). Endoreduplication medierad av den anafasfrämjande komplexa aktivatorn CCS52A krävs för symbiotisk celldifferentiering i Medicago truncatula-knölar. Växtcell 15, 2093-2105. doi: 10.1105/tpc.014373
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Walker, S. A. och Downie, J. A. (2000). Inträde av Rhizobium leguminosarum bv. viciae i rothår kräver minimal Nodfaktorspecificitet, men efterföljande infektionstrådtillväxt kräver nodO eller nod. Mol. Växtmikroben Interagerar.13, 54–62. doi: 10.1094/MPMI.2000.13.7.754
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text