- introduktion
- material och metoder
- växtmaterial och Tillväxttillstånd
- psi-och PSII-mätningar
- Elektrokromisk Skiftanalys
- Fotoinhibitoriska behandlingar
- statistisk analys
- resultat
- diskussion
- rollen för CEF-stimulering vid svagt ljus
- förändring i P700-Oxidationsförhållandet vid svagt ljus är inte relaterat till pmf
- Etrii kontrollerar P700-Oxidationsförhållandet vid svagt ljus
- slutsats
- Författarbidrag
- finansiering
- intressekonflikt uttalande
introduktion
även om ljus är drivkraften för fotosyntes kan högt ljus orsaka betydande fotoinhibition av fotosystem II (PSII) för blad av skuggbildande växter (Kitao et al., 2000; Barth et al., 2001; Krause et al., 2004; Huang et al., 2015b, 2016b, c). Photoinhibited PSII återhämtar sig dock snabbt vid svagt ljus (He och Chow, 2003; Zhang och Scheller, 2004, Allakhverdiev et al., 2005), på grund av den snabba omsättningshastigheten för D1-protein (Aro et al., 1993; Zhang och Scheller, 2004; Allakhverdiev et al., 2005). Hastigheten för PSII-reparation reducerades vid hämning av syntesen av ATP antingen via PSI eller PSII (Allakhverdiev et al., 2005), vilket indikerar att den snabba reparationen av fotodamaged PSII-komplex behöver en stor mängd ATP på kort tid. En tidigare studie visade att efter kylinducerad fotoinhibition av PSII stimulerades cykliskt elektronflöde (CEF) signifikant under återhämtning vid svagt ljus (Huang et al., 2010). Det föreslogs att denna stimulering av CEF vid svagt ljus förbättrade syntesen av ATP för snabb reparation av PSII (Huang et al., 2010). Men mer bevis behövs för att klargöra den specifika rollen för CEF-stimulering vid svagt ljus efter PSII-fotoinhibition.
under CEF cyklas elektroner från antingen NADPH eller ferredoxin runt PSI i plastokinonpoolen. Denna elektronöverföring är kopplad till protontranslokation och genererar en protongradient över thylakoidmembran (Bisexuell) (Shikanai och Yamamoto, 2017). Förutom att bidra till ATP-syntes, är en annan funktion av sackaros nedreglering av fotosyntetisk elektrontransport genom att surgöra thylakoidlumen (Shikanai, 2014, 2016). Denna reglering innefattar två olika mekanismer: en är kopplad till termisk energiavledning och sprider överskott av absorberad ljusenergi som värme från PSII-antenner (Takahashi et al., 2009), och den andra är nedreglering av Cyt b6/f-komplex aktivitet och kontrollerar elektronöverföringshastigheten till PSI (Suorsa et al., 2012, 2016; Tikkanen och Aro, 2014). I angiospermer inklusive Arabidopsis thaliana fungerar två vägar för PSI-cyklisk elektrontransport (Shikanai, 2007). Den första CEF-vägen är PGR5- / PGRL1-beroende väg känslig för antimycin A (Munekage et al., 2002; Sugimoto et al., 2013), och den andra är beroende av kloroplast NADH dehydrogenasliknande (NDH) komplex (Burrows et al., 1998; Shikanai et al., 1998). Bidraget från den PGR5-/PGRL1-beroende vägen är mer signifikant i C3-växter. Under högt ljus inducerar aktiveringen av PGR5-/PGRL1-beroende CEF försurningen av tylakoidlumen och leder således till de höga nivåerna av P700-oxidationsförhållande (Suorsa et al., 2012, 2016; Kono et al., 2014; Yamori et al., 2016). Som jämförelse påverkades P700-oxidationsförhållandet vid svagt ljus lite av bristen i PGR5-/PGRL1-beroende CEF (Munekage et al., 2002, 2004; Kono et al., 2014). Vid måttlig PSII-fotoinhibition åtföljdes stimuleringen av CEF vid svagt ljus med en ökning av P700-oxidationsrationen (Huang et al., 2010). Det är emellertid oklart om detta ökade P700-oxidationsförhållande orsakas av CEF-stimuleringen.
förutom CEF spelar elektronöverföring från PSII (ETRII) en viktig roll för att påverka redoxtillståndet för PSI (Tikkanen et al., 2014; Huang et al., 2016a, c; Suorsa et al., 2016). I pgr5-växter av A. thaliana kommer allvarlig fotoinhibition av PSII sannolikt att fungera som den ultimata kontrollen av fotosyntetisk elektronöverföring som möjliggör underhåll av P700 optimalt oxiderad under överskott av ljus (Tikkanen et al., 2014). Nyligen, Suorsa et al. (2016) indikerade att pgr5-växter visade högt P700-oxidationsförhållande under högt ljus när ETRII reducerades genom att kombinera knockout-mutationer i PsbO1, PsbP2, PsbQ1, PsbQ2 och PsbR loci. I kylda tobaksblad deprimerade måttlig PSII-fotoinhibition elektronflödet till PSI och ökade sedan P700-oxidationsförhållandet under ytterligare kylningsbehandlingar (Huang et al., 2016a). Vidare ledde kylinducerad fotoinhibition av PSII till en depression av ETRII, vilket åtföljdes av en ökning av P700-oxidationsförhållandet vid svagt ljus (Huang et al., 2010). Vi antar att P700-oxidationsförhållandet vid svagt ljus styrs främst av elektronflöde från PSII.
i vår tidigare rapport observerade vi att den skuggbildande växten Panax notoginseng visade selektiv fotoinhibition av PSII under högljusspänning. Efter kortvarig exponering för högljusspänning minskade ETRII vid svagt ljus signifikant men CEF stimulerades signifikant. Samtidigt ökade P700-oxidationsförhållandet i stor utsträckning. Våra specifika mål var att (1) undersöka om CEF-stimulering vid svagt ljus huvudsakligen underlättar syntesen av ATP; och (2) Bestäm om ökningen av P700-oxidationsförhållandet vid måttlig PSII-fotoinhibition är mer relaterad till ETRII snarare än CEF-stimuleringen eller förändringen i pmf. För att ta itu med dessa frågor behandlades intakta blad av Panax notoginseng vid 2258 kcal fotoner m-2 s-1 i 30 minuter. Före och efter högljusbehandling bestämdes energifördelningen i PSII, redoxtillståndet för PSI och protonmotivkraft (pmf) vid ett svagt ljus på 54 occylmol fotoner m-2 s-1.
material och metoder
växtmaterial och Tillväxttillstånd
i den aktuella studien användes 2-åriga växter av en skuggbildande växt Panax notoginseng (Burkill) F. H. Chen ex C. Chow och W. G. Huang för experiment. Dessa växter odlades i lätt tillstånd av 10% solljus (med maximal ljusintensitet på cirka 200 occurmol fotoner m-2 s-1). Inga vatten-och näringsbelastningar upplevdes för dessa växter. 9 veckor gamla helt expanderade blad användes för fotosyntetiska mätningar.
psi-och PSII-mätningar
fotosystem I-och PSII-parametrar mättes vid 25 CCB genom att samtidigt registreras med hjälp av ett dubbelt Pam-100-mätsystem (Heinz Walz GmbH, Effeltrich, Tyskland). Klorofyllfluorescensparametrarna beräknades enligt följande: Fv / Fm = (Fm-Fo)/Fm, Y(II) = (Fm’ – Fs) / Fm’ (Genty et al., 1989), NPQ = (Fm-Fm’) / Fm’. Fo och Fm är den minsta och maximala fluorescensen efter mörk acklimatisering. Fs är den ljusanpassade steady-state fluorescensen. Fo och Fm bestämdes efter mörk acklimatisering i 30 minuter före och efter högljusbehandling. PSI-fotosyntetiska parametrar mättes med dubbla PAM-100 baserat på P700-signal (skillnad i intensiteter på 830 och 875 nm pulsmodulerat mätljus som når fotodetektorn) (Klughammer och Schreiber, 2008). P700 + – signalerna (P) kan variera mellan en minimal (P700 helt reducerad) och en maximal nivå (P700 helt oxiderad). Den maximala nivån (Pm) bestämdes med applicering av en mättnadspuls (600 ms och 10000 occurmol fotoner m-2 s-1) efter förbelysning med långt rött ljus och Pm användes för att uppskatta PSI-aktiviteten. Pm ’ bestämdes liknande Pm men med aktiniskt ljus istället för långt rött ljus. Kvantutbytet av PSI beräknades som Y (I) = (Pm’- P)/Pm. P700-oxidationsförhållandet i ett givet aktiniskt ljus beräknades som Y (ND) = P/Pm. Kvantutbytet av PSI icke-fotokemisk energiavledning på grund av acceptorsidan begränsning beräknades som Y (NA) = (Pm-Pm’)/Pm. Steady state-värdena för Y(II), NPQ, Y(i) och Y (ND) mättes efter lätt acklimatisering vid svagt ljus i 20 minuter.
Fotosyntetiska elektron flöde genom PSI och PSII har beräknats som: ETRII = Y (II) ×PPFD ×0.84 ×0.5 (Krall och Edwards, 1992), ETRI = Y (I) ×PPFD×0.84 ×0.5 (Yamori et al., 2011), där 0,5 antas vara andelen absorberat ljus som når PSI eller PSII, och 0,84 antas vara absorptansen (fraktionen av det infallande ljuset som absorberas av löv). Omfattningen av CEF-aktivering uppskattades som ETRI/ETRII-förhållande (Yamori et al., 2011, 2015). Det bör noteras att 1: 1 excitationspartitionering mellan PSI och PSII kanske inte är verklig, eftersom skuggväxter tenderar att öka PSII/PSI-förhållandet (Lunde et al., 2003; Tikkanen et al., 2006; Grieco et al., 2012). Vidare mättes i denna studie psi-och PSII-parametrar med rött aktiniskt ljus (635 nm) tillhandahållet av Dual-PAM 100 (Walz, Tyskland). Eftersom rött aktiniskt ljus gynnar exciteringen av PSII över PSI, kan exciteringsenergifördelningen från LHCII till PSII och PSI påverkas av det röda aktiniska ljuset (Tikkanen et al., 2017).
Elektrokromisk Skiftanalys
ECS-signalen övervakades som absorbansförändringen vid 515 nm med användning av en Dual-PAM-100 (Walz, Effeltrich, Tyskland) utrustad med en p515/535 emitter-detektormodul (Walz). ECS-signalen erhölls efter 20 min belysning vid 54 occumol fotoner m-2 s-1, Därefter mättes ECS-sönderfallet genom att stänga av det aktiniska ljuset i 30 s. analysen av ECS dark interval relaxation kinetics (DIRKECS) gjordes med metoden för Sacksteder et al. (2001) och Takizawa et al. (2008). Total pmf uppskattades från den totala amplituden för det snabba förfallet av ECS-signalen under 300 ms mörkt intervall. Den långsamma avslappningen av ECS-signalen möjliggjorde att känna igen bidraget från protongradient över thylakoidmembranen (Askorph). Tidskonstanten för första ordningens ECS-avkoppling (tECS) är omvänt proportionell mot protonledningsförmågan (gH+) hos tylakoidmembranet genom ATP-syntas (Sacksteder och Kramer, 2000; Cruz et al., 2005). Som ett resultat uppskattades gH+ som inversen av sönderfallstidskonstanten .
Fotoinhibitoriska behandlingar
efter mörk acklimatisering i 30 minuter mättes Fv/Fm och Pm i intakta blad. Efteråt var dessa intakta blad ljusanpassade vid 59 occylmol fotoner m – 2 s-1 för 20 min och parametrar för klorofyllfluorescens, P700-signal och ECS-signal registrerades. Därefter ändrades det aktiniska ljuset till 2258 oc-mmol fotoner m-2 s-1. Efter exponering för detta höga ljus i 30 minuter ändrades det aktiniska ljuset till 59 occylmol fotoner m-2 s-1 omedelbart och de fotosyntetiska parametrarna registrerades efter lätt acklimatisering i 20 minuter. Slutligen mättes Fv / Fm och Pm efter mörk acklimatisering i 30 minuter.
statistisk analys
resultaten visades som medelvärden för fem oberoende experiment. Oberoende t-test användes vid 0,05 signifikansnivå för att avgöra om det fanns signifikanta skillnader mellan olika behandlingar. Alla statistiska analyser utfördes med hjälp av SPSS 16.0.
resultat
under fotosyntetisk induktion vid 59 occolmol fotoner m – 2 s-1 ökade etrii gradvis och nådde ett steady state vid cirka 18 min (Figur 1a). Som jämförelse var ETRI konstant under denna induktionsfas (Figur 1a). Efter uppkomsten av detta låga ljus var ETRI mycket högre än ETRII. Men efter en 20-min fotosyntetisk induktion var ETRI lägre än ETRII. Dessa resultat föreslog aktivering av CEF under den inledande fasen av induktion, i enlighet med tidigare studier (Joliot och Joliot, 2002, 2005; Makino et al., 2002). Efter denna fotosyntetiska induktion vid svagt ljus, bladen belystes vid ett högt ljus av 2258 occurmol fotoner m – 2 s-1 för 30 min. Intressant minskade ETRII gradvis under högljusbehandlingen (Figur 1b). Under tiden var värdet av ETRI högre än det för ETRII (Figur 1b), vilket indikerar aktiveringen av CEF under högt ljus. Efter denna högljusbehandling bibehölls den maximala fotooxiderbara P700 (Pm) stabil (figur 2a). Som jämförelse minskade den maximala fluorescensintensiteten (Fm) med 40% och det maximala kvantutbytet av PSII (Fv/Fm) minskade från 0,80 till 0,65 (figurerna 2b,C). Dessa resultat indikerade den selektiva fotoinhibitionen av PSII i blad av Panax notoginseng under högljusspänning.
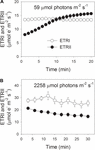
figur 1. Fotosyntetisk elektronflöde vid svagt ljus (A) och högt ljus (B). (A) mörk-acklimatiserade blad utsattes för ett svagt ljus av 59 oc-mmol fotoner m-2 s-1 för 20 min. (B) efter belysning vid 59 occolmol fotoner m-2 s-1 för 20 min för att aktivera elektronsänken i fotosyntesen exponerades bladen för ett högt ljus på 2258 occolmol fotoner m-2 s-1 för 30 min. Värdena är medel för att beteckna det som är nödvändigt för att beteckna det som är nödvändigt.
före högljusbehandling upplystes mogna löv vid ett svagt ljus av 59 occbmol fotoner m – 2 s-1 i 20 minuter för att aktivera fotosyntes, och sedan registrerades värden för ETRI, ETRII, Y(ND) och NPQ. Efter högljusbehandling i 30 min registrerades värden för ETRI, ETRII, Y(ND) och NPQ efter en ny acklimatiseringsfas på 20 min till svagt ljus. Före högljusbehandling var värdena för ETRI och ETRII vid 59 oktmolfotoner m-2 s-1 13,4 respektive 15,7 oktmolelektroner m-2 s-1 (figurerna 3A, B). Efter högljusbehandling var ETRI och ETRII vid svagt ljus 12,8 respektive 11,2 kubolelektroner m-2 s-1 (figurerna 3A, B). Värdet av ETRII vid 59 oc-mmol fotoner m – 2 s-1 minskade med cirka 30% efter fotoinhibitorisk behandling, vilket indikerar depression av ETRII vid måttlig PSII-fotoinhibition. Före högljusbehandling var värdet av ETRI / ETRII-förhållandet vid 59 oc-molfotoner m-2 s-1 0,85 (figur 3C). Efter fotoinhibitorisk behandling ökade etri / ETRII-förhållandet signifikant till 1,14 (figur 3C). Dessa resultat föreslog stimulering av CEF vid svagt ljus vid PSII-fotoinhibition (Yamori et al., 2011, 2015).
efter högljusbehandlingen ändrades inte steady-state-värdet för Y(i) vid 59 occylmol fotoner m-2 s-1 (figur 4a). Intressant nog minskade Y(NA) signifikant från 0,36 till 0,23 (figur 4B) och Y(ND) ökade signifikant från 0,1 till 0,25 (figur 4c). Dessa resultat indikerade förändringen i redoxtillstånd för PSI vid svagt ljus efter fotoinhibitorisk behandling. Under tiden ändrades värdet på NPQ något (figur 4D). Som ett resultat hade den måttliga PSII-fotoinhibitionen olika effekter på P700-oxidationsförhållandet och NPQ vid svagt ljus.
för att klargöra om ökningen av Y(nd) orsakas av en ökning av pmf, bestämdes de elektrokroma skiftsignalerna vid 54 kazakmol fotoner m-2 s-1 Efter lätt acklimatisering i 20 minuter före och efter högljusbehandlingen. Intressant nog minskade den totala pmf signifikant med 18% Efter högljusbehandling (figur 5A), men nivån på 0bph förändrades obetydligt (figur 5B). Eftersom bildandet av pmf kan påverkas av thylakoidprotonledningsförmågan undersöktes också protonledningsförmågan (gH+) hos thylakoidmembranet vid detta låga ljus före och efter högljusbehandling. Resultatet visade att gH + inte förändrades med högljusbehandlingen (figur 5C), vilket tyder på att högljusbehandlingen knappast påverkade aktiviteten hos kloroplast ATP-syntas vid svagt ljus. Eftersom minskningen av 30% i ETRII åtföljdes av en minskning med 18% i pmf, kompenserade stimuleringen av CEF vid svagt ljus delvis för bildandet av pmf och struphuvud.

figur 5. Värden för den totala protonmotivkraften (pmf) (a), protongradienten (sackaros) över tylakoidmembranen (B) och protonledningsförmågan (gH+) hos tylakoidmembranet (C) vid svagt ljus. Alla parametrar mättes efter anpassning vid 54 kcal fotoner m – 2 s-1 för 20 min. Värdena är medel för att beteckna det som är nödvändigt för att beteckna det som är nödvändigt. Asterisk indikerar en signifikant (P < 0.05) förändring efter högljusbehandling.
diskussion
rollen för CEF-stimulering vid svagt ljus
det har indikerats att CEF spelar viktiga roller för att upprätthålla fotosyntes och växttillväxt (Yamori och Shikanai, 2016). Under högt ljus anses CEF nu vara avgörande för att balansera ATP/NADPH-energibudgeten samt för att skydda PSI och PSII från skador (Munekage et al., 2002, 2004; Takahashi et al., 2009; Suorsa et al., 2012, 2016; Walker et al., 2014; Huang et al., 2015a, 2017b). Vid låg ljusintensitet spelar CEF en viktig roll för att optimera fotosyntetisk CO2-assimilering troligen via tillförsel av extra ATP (Yamori et al., 2011, 2015; Nishikawa et al., 2012; Huang et al., 2015a). CEF: s huvudroll moduleras flexibelt som svar på ljusintensiteten. I synnerhet bidrar CEF huvudsakligen till att balansera ATP / NADPH-energibudgeten under submättande ljusintensiteter, men skyddar främst fotosyntetiska apparater mot fotoinhibition när de utsätts för mättande ljusintensiteter via försurning av thylakoidlumen (Huang et al., 2015a). Intressant, Huang et al. (2010) fann att CEF stimulerades signifikant vid svagt ljus efter kylinducerad fotoinhibition av PSII och antog att denna CEF-stimulering huvudsakligen förbättrade syntesen av ATP för snabb reparation av PSII. Men mer bevis behövdes för att stödja denna hypotes. I den aktuella studien observerade vi att värdet av ETRI/ETRII vid svagt ljus ökade signifikant (figur 3C), vilket indikerar stimulering av CEF runt PSI (Huang et al., 2011, 2012, 2017a, B; Yamori et al., 2011, 2015). Samtidigt minskade amplituden för pmf signifikant och Accordph minskade något. Dessa resultat indikerade att stimuleringen av CEF inte utlöste den nigerianska beroende nedregleringen av fotosyntetisk elektrontransport. Med andra ord underlättade denna stimulering av CEF vid svagt ljus huvudsakligen syntesen av ATP.
även om protonkonduktiviteten hos kloroplast ATP-syntas inte påverkades av högljusbehandlingen resulterade Den mindre storleken på pmf i en minskning av den faktiska hastigheten för ATP-syntes. Hastigheterna för PSII-reparation och fotosyntetisk CO2-assimilering vid svagt ljus begränsas huvudsakligen av ATP-synteshastigheten (Allakhverdiev et al., 2005; Yamori et al., 2011, 2015; Nishikawa et al., 2012). Eftersom ETRII minskade signifikant vid PSII-fotoinhibition reducerades hastigheten för ATP-syntes via ETRII anmärkningsvärt. För att balansera ATP / NADPH-förhållandet som krävs av primär metabolism och snabb reparation av PSII behövs en flexibel mekanism för att ge extra ATP. Under detta tillstånd minskade ETRII med 30% men den totala pmf minskade med bara 18%. Dessa resultat indikerar att stimuleringen av CEF kompenserade för minskningen av ETRII-beroende bildning av pmf, vilket förbättrar syntesen av ATP. Den snabba reparationen av fotodamaged PSII är beroende av ATP-syntes (Allakhverdiev et al., 2005). Acceleration av CEF runt PSI ökade den intracellulära koncentrationen av ATP, vilket accelererade hastigheten för PSII-reparation i Synechocystis (Allakhverdiev et al., 2005). Sammantaget föreslår vi att stimuleringen av CEF vid svagt ljus spelar en viktig roll i den snabba reparationen av PSII-aktivitet via en ytterligare ATP-syntes.
förändring i P700-Oxidationsförhållandet vid svagt ljus är inte relaterat till pmf
ett intressant fenomen är att P700-oxidationsförhållandet vid svagt ljus ökade signifikant vid måttlig PSII-fotoinhibition (Huang et al., 2010), vilket också visas i den aktuella studien. Under förhållanden där absorberat ljus överstiger kraven för fotosyntes reglerar CEF-beroende generering av sackaros ned aktiviteten för Cyt b6/f-komplex aktivitet och kontrollerar elektronflödet från PSII till PSI (Suorsa et al., 2012, 2016; Shikanai, 2014, 2016), optimera redoxtillståndet för P700 i PSI och minimera ROS-produktion i PSI under fotosyntes. I pgr5-växter av A. thaliana reduceras P700 med elektroner i ljuset på grund av förlusten av PMF-generation. I det pgr5-mutanta ackumulerande flavodiironproteinet återställdes emellertid PMF-nivån till vildtypsnivån och sålunda oxiderades P700 som i vildtypen (Yamamoto et al., 2016). I klorofyll b-bristande vetemutantlinjer leder otillräcklig tylakoidprotongradient till överreduktion av PSI-acceptorsidan och därmed PSI-fotoinhibition under högt ljus eller hög temperatur (Brestic et al., 2015, 2016). Nyligen rapporterade vissa studier att den ökade aktiviteten hos kloroplast ATP-syntas försämrade bildandet av pmf och orsakade överreduktionen av fotosyntetisk elektrontransportkedja, vilket resulterade i fotodamage av PSI under högt ljus och fluktuerande ljus (Kanazawa et al., 2017; Takagi et al., 2017). Dessa rapporter stöder PMF: s kritiska roll för att upprätthålla P700 optimalt oxiderad under överskott av ljusenergi.
våra nuvarande resultat indikerade att efter högljusbehandlingen i 30 minuter minskade den totala pmf som bildades vid detta låga ljus signifikant och bildandet av Ubisph förändrades något (figurerna 5A,B). Under tiden ökade P700-oxidationsförhållandet signifikant från 0,1 (före behandling) till 0,25 (efter behandling) (figur 4C). Dessa resultat indikerar starkt att denna ökning av P700-oxidationsförhållandet inte kan förklaras av förändringen i pmf, vilket i stor utsträckning skiljer sig från korrelationen mellan Y(ND) och pmf under högt ljus (Yamamoto et al., 2016; Takagi et al., 2017). Därför är PMF: s reglerande effekt på P700-oxidationsförhållandet minimal vid svagt ljus men blir särskilt avgörande under högt ljus eller fluktuerande ljus. Under högt ljus åtföljs vanligtvis ett högt värde av Y(ND) med ett högt värde av NPQ (Munekage et al., 2002, 2004; Kono et al., 2014; Zivcak et al., 2015), på grund av ökningen av pmf. Försämringen av bildandet av usci leder till minskade nivåer av Y (ND) och NPQ under högt ljus (Suorsa et al., 2012; Kono et al., 2014; Yamamoto et al., 2016; Kanazawa et al., 2017). Induktionen av NPQ vid svagt ljus bestäms huvudsakligen av nivån av lumenförsurning. Efter högljusbehandling förändrades inte omfattningen av lumensyrning (figur 5B) och NPQ förblev stabil (figur 4D). Som jämförelse ökade Y (ND) signifikant. Dessa resultat indikerar att vid svagt ljus kontrollerades Y(ND) och NPQ av olika regleringsmekanismer.
Etrii kontrollerar P700-Oxidationsförhållandet vid svagt ljus
PSI blir överreducerat endast när elektronflödet från PSII överstiger kapaciteten hos PSI-elektronacceptorer för att klara elektronerna. När elektronflödet till PSI är begränsat är PSI extremt tolerant mot ljusspänning i pgr5-växterna i A. thaliana (Suorsa et al., 2012, 2016; Tikkanen et al., 2014). För den skuggbildande växtpsykotria rubra var PSI-aktivitet okänslig för högljusspänning i närvaro av DCMU (Huang et al., 2016c). I kylda tobaksblad möjliggjorde måttlig PSII-fotoinhibition underhållet av P700 optimalt oxiderad och sedan skyddad PSI-aktivitet mot ytterligare fotodamage (Huang et al., 2016a). Dessa resultat indikerade etriis viktiga roll för att kontrollera redoxtillståndet för PSI under höga ljus-eller kylningsljusspänningar. Våra nuvarande resultat visade att den måttliga PSII-fotoinhibitionen ledde till en signifikant depression av ETRII vid svagt ljus. Under tiden förändrades PSI-aktiviteten och aktiviteten hos kloroplast-ATP-syntas något, och den totala pmf-och struphuvud ökade inte. Som ett resultat orsakades inte reduktionen i ETRII av den beroende fotosyntetiska kontrollen via CYT b6/f-komplexet utan inducerades troligen av minskningen av PSII-aktivitet (Tikkanen et al., 2014). Den minskade tillförseln av elektroner från PSII till PSI ledde till den ökade nivån av P700-oxidation. Således styrdes P700-oxidationsförhållandet vid svagt ljus till stor del av ETRII.
slutsats
Vi fann att den selektiva fotoinhibitionen av PSII inducerade en stimulering av CEF och en ökning av P700-oxidationsförhållandet vid svagt ljus. Stimuleringen av CEF utlöste inte den beroende Nedregleringen av den fotosyntetiska elektrontransporten i enlighet med Uisph. Som ett resultat underlättade denna stimulering av CEF vid svagt ljus huvudsakligen syntesen av ATP, vilket är väsentligt för snabb reparation av fotodamaged PSII. Ökningen i P700-oxidationsförhållandet kunde inte förklaras av förändringen i den beroende fotosyntetiska kontrollen i enlighet med den beroende fotosyntesen i CYT B6/f-komplexet, men orsakades främst av den minskade tillförseln av elektroner från PSII till PSI.
Författarbidrag
WH, S-BZ och TL utformade och utformade forskningen. WH och Y-JY genomförde experimenten. WH, Y-JY, S-BZ och TL analyserade data. WH skrev det första utkastet till manuskriptet, som intensivt redigerades av alla författare.
finansiering
denna studie stöddes av National Natural Science Foundation of China (31670343), Youth Innovation Promotion Association of the Chinese Academy of Sciences (2016347) och de stora Vetenskaps-och Teknikprogrammen i Yunnan-provinsen (2016zf001).
intressekonflikt uttalande
författarna förklarar att forskningen genomfördes i avsaknad av kommersiella eller finansiella relationer som kan tolkas som en potentiell intressekonflikt.
Kono, M., Noguchi, K. och Terashima, I. (2014). Roller av det cykliska elektronflödet runt PSI (CEF-PSI) och O2-beroende alternativa vägar i reglering av det fotosyntetiska elektronflödet i kortsiktigt fluktuerande ljus i Arabidopsis thaliana. Växtcell Physiol. 55, 990–1004. doi: 10.1093 / pcp / pcu033
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Shikanai, T. (2016). Reglerande nätverk av protonmotivkraft: bidrag från cyklisk elektrontransport runt fotosystem I. Photosynth. Res 129, 253-260. doi: 10.1007 / s11120-016-0227-0
PubMed Abstract / CrossRef fulltext / Google Scholar
Yamori, W., Makino, A. och Shikanai, T. (2016). En fysiologik roll för cyklisk elektrontransport runt fotosystem är för att upprätthålla fotosyntes under fluktuerande ljus i ris. Sci. Rep. 6: 20147. doi: 10.1038 / srep20147
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Yamori, W. och Shikanai, T. (2016). Fysiologiska funktioner för cyklisk elektrontransport runt fotosystem I för att upprätthålla fotosyntes och växttillväxt. Annu. Rev. Växt Biol. 67, 81–106. doi: 10.1146 / annurev-arplant-043015-112002
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Yamori, W., Shikanai, T. och Makino, A. (2015). Fotosystem I cykliskt elektronflöde via kloroplast NADH-DEHYDROGENASLIKNANDE komplex utför en fysiologisk roll för fotosyntes vid svagt ljus. Sci. Rep. 5: 13908. doi: 10.1038 / srep13908
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Zhang, SP och Scheller, HV (2004). Photoinhibition av photosystem I vid kylning temperatur och efterföljande återhämtning i Arabidopsis thaliana. Växtcell Physiol. 45, 1595–1602. doi: 10.1093 / pcp / pch180
PubMed Abstrakt / CrossRef fulltext / Google Scholar