A Microbial Biorealm page on the genus Lactobacillus delbrueckii
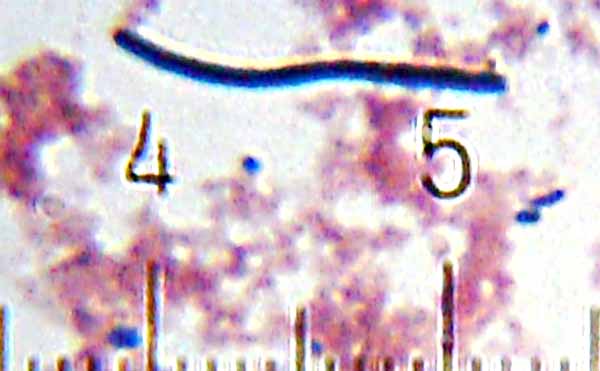
Numbered ticks are 11 µM apart.
Gram-stained.
Photograph by Bob Blaylock.
Classification
Higher order taxa
Bacteria (Domain); Firmicutes (Phylum); Bacilli (Class); Lactobacillales (Order); Lactobacillaceae (heimo)
laji
NCBI: taksonomia
Lactobacillus delbrueckii
subspecies: bulgaricus, lactis, delbrueckii, and indicus
kuvaus ja merkitys
Lactobacillus us delbrueckii on sauvan muotoinen, grampositiivinen, liikkumaton bakteeri. Lajille on yhteistä se, että se pystyy hapattamaan sokerialustoja maitohappotuotteiksi anaerobisissa olosuhteissa. Sellaisenaan L. delbrueckii esiintyy yleensä maitotuotteissa, kuten jogurtissa, maidossa ja juustossa L. delbrueckii subsp: tä lukuun ottamatta. delbruecki, jotka sijaitsevat vihannesten lähteissä (3). Lajista tunnetaan tähän mennessä neljä alalajia, jotka eroavat toisistaan sen aineenvaihduntatuotteiden ja sisäisen genetiikan perusteella. Viimeisin hyväksytty alalaji, L. delbrueckii subsp. indicus eristettiin Intialaisesta meijeristä (1). Sen sijaan tohtori Stamen Grigorov eristi L. delbrueckii subsp: n. bulgaricus jogurttinäytteestä vuodelta 1905.
ominaisuudet, jotka määrittelevät L. delbrueckii: n homofermentiiviseksi maitohappobakteeriksi (LAB), eivät rajoitu sen metaboliseen lopputuotteeseen D-laktaattiin ja L-laktaattiin. L. delbrueckii subsp. bulgaricuksella on osoitettu olevan probioottisia vaikutuksia ihmisiin ja eläimiin, joihin kuuluu laktoosin parempi sietokyky ja kyky stimuloida immuunivastetta (4, 5, 7). Aiemmat väittelyt tätä tietoa vastaan kyseenalaistivat jälkimmäisen kyvyn selviytyä matalissa happamissa ympäristöissä ja ihmisen ruoansulatuskanavan mahanesteissä. Fosfopolysakkaridi, jota tuottaa L. delbrueckii subsp. bulgaricus kykenee tehostamaan makrofagien fagosytoosia hiirillä (4).
genomin rakenne
Lactobacillus delbrueckii subsp: n pyöreä genomi. bugaricus ATCC 11842 valmistui toukokuussa 2006. Se koostuu 1 864 998 nukleotidista, ja sen G-C-pitoisuus on epätavallisen suuri (49%) verrattuna muihin laktobasilli-suvun lajeihin, joihin se kuuluu. 2 217 geenistä 1 562 koodaa proteiineja ja 533 pseudogeenejä (10). Tämänkaltaiset genomiset piirteet sekä lisäyssekvenssimallit osoittavat sen sopeutumista meijeriteollisuudessa ja tukevat teoriaa nopeasta evoluutiovaiheesta (11).
proteiineja koodaavista 1 562 geenistä prtB ja lac operoni ovat tärkeitä L. delbrueckiin homofermentatiivisille ominaisuuksille. Sisällä lac operon on lacS, lacZ, ja lacR geenit, jotka koodaavat ottamista ja jakautuminen laktoosin.(3) Lacs-geeni koodaa laktoosiläpäisyä, joka vastaa laktoosin kuljetuskyvystä kalvon läpi. Laktoosin aineenvaihdunnalle välttämätön tärkeä entsyymi B-galaktosidaasi koodataan lacZ-geeniin. Laczin alajuoksulla on säätelygeeni lacR.
solujen rakenne ja aineenvaihdunta
grampositiivisena bakteerina L. delbrueckii säilyttää violetin tahransa Gram-testissä. Tämäntyyppisille mikrobeille on ominaista paksu soluseinä ja solukalvo. Ylimääräisenä esteenä toimivan ulomman kalvon puuttuminen voi olla syynä sen herkkyyteen bakteriofagihyökkäyksille (2).
prtB-geenin koodaamia proteaaseja tavataan ankkuroituneena L. delbrueckii subsp: n soluseinämään. bulgaricus ja lactis; ja todennäköisesti subsp. indicus. Alalajien kyky kasvaa maidossa johtuu kaseiinin hajoamisessa tapahtuvasta entsymaattisesta aktiivisuudesta, joka altistaa välttämättömät aminohapot lisäksi lacz-geenin konstitutiiviselle tai indusoituvalle ilmentymiselle. (8,4)
merkitsevää neljälle L. delbrueckii-alalajille on sen metaboloimien substraattien määrä ja tyypit. Kuten todettiin, tällaiset ominaisuudet rajoittuvat entsyymin ilmentymiseen sen genomissa. L. delbrueckii subsp. bulgaricus ja subsp. indicus voi metaboloida laktoosia, glukoosia, fruktoosia ja mannoosia. Näiden lisäksi L. delbrueckii subsp. lactis katabolisoi galaktoosia, sakkaroosia, maltoosia, trehaloosia ja muita muunneltuja hiilihydraatteja.(4)
ekologia
hapatettujen maitotuotteiden asukkaana ja maitohapon tuottajana Lactobacillus delbrueckii lukuun ottamatta L. subsp: tä. delbrueckii, aiheuttaa sen alhaisen happaman ympäristön. Ravinnontarve on mukautettu bakteerin ympäristöön; se sisältää muun muassa aminohappoja, vitamiineja, hiilihydraatteja ja tyydyttymättömiä rasvahappoja (9). L. delbrueckiin optimaalinen kasvulämpötila on 40-44 °C anaerobisissa olosuhteissa(3). Erityisesti L. subsp. bulgaricuksella on symbioottinen suhde Streptococcus thermophilus-bakteerin kanssa, koska se elää rinnakkain maitohappobakteeriviljelmissä.
patologia
Lactobacillus delbrueckii on ei-patogeeninen. Sitä käytetäänkin laajalti elintarviketeollisuudessa, ja sitä voidaan käyttää jogurteissa, maidoissa, vihanneksissa ja juustoissa.
soveltaminen biotekniikkaan
neljästä tähän mennessä tunnetusta alalajista L. delbrueckii subsp. bulgaricus ja subsp. maitoteollisuudelle lactit ovat tärkeimpiä, sillä ne ovat käyneen maidon, jogurtin ja juuston valmistuksen aloitusviljelmiä. Taloudelliset menetykset olisivat merkittäviä, jos laajalti käytetyn Lactobacillus delbrueckii subsp: n käymisprosessi olisi käynnissä. bulgaricus ja subsp. lactis estettiin. Näin ollen meijeriteollisuuden on kyettävä havaitsemaan bakteriofagit ja sopeuttamaan tuotanto-olosuhteita turvallisuuden ja säilyvyyden korkean laadun varmistamiseksi (2). Johtuen L. delbrueckii subsp: n symbioottisesta suhteesta. bulgaricus ja Streptococcus thermophilus on tutkittu samanaikaisesti.
tämänhetkinen tutkimus
laktobasillien spesifisillä kannoilla on osoitettu olevan mitogeenisiä vaikutuksia ja apua pernan solujen proliferaatiossa. L. delbrueckii-alalajien bulgaricus ja L. acidophilus lämpökäsitellyt YS-kannat aiheuttivat suoraan IgM: n ja IgG: n muodostumisen hiiren spenosyyteissä ja olivat riippuvaisia bakteeripitoisuuksista toisin kuin ATCC-kannat. Ensin mainittu oli tehokkain määrillä 5×106 ja 2 × 107 laktobasilli ml–1. Vasta-ainepitoisuudet määritettiin Elisan ja Fisherin testillä. Lisäksi molempien Laktobasillilajien kannat YS ja ATCC aiheuttivat lymfosyyttien proliferaatiota. L. delbrueckii subsp. bulgaricuksen on osoitettu aktivoivan polyklonaalisia B-soluja, jotka on osoitettu ylläpitämällä korkeita vasta-ainepitoisuuksia laktobasillien vasta-aineiden poistamisen jälkeen. (13)
eräässä tutkimuksessa ehdotettiin multiplex PCR: n käyttöä bakteriofagien havaitsemiseen missä tahansa valmistusvaiheessa. Menetelmä osoittautui yksinkertaiseksi ja nopeaksi, ja samalla taattiin tuotteiden vähimmäislaatuvaatimukset. Vaikka tulokset osoittivat pieniä määriä L. delbrueckii-fageja käytetyistä näytteistä löytyi suhteellisesti suurempi määrä S. thermophilus-fageja. Nämä tulokset johtuvat siitä, että S. thermophilus-bakteeria käytetään yhä enemmän aloitusviljelmissä. (2)
Lactobacillus delbrueckii ei yleensä esiinny maitoteollisuuden aloitusviljelmien ulkopuolella. Luonnollista ympäristöä, josta se on peräisin, ei tiedetä varmasti. Tuore tutkimus kertoi L. delbrueckii subsp: n eristyneisyydestä ja luonnehdinnasta. bulgaricus yhdessä sen symbiontti Streptococcus thermophilus kasveista Bulgariassa perusteella perinteisen jogurtti valmistelu. Kuusisataakuusikymmentäviisi kasvinäytettä, joiden kohdekasvi Cornus mas kerättiin neljältä paikalta pois Ihmisasutuksen läheisyydestä. L. subsp: n tunnistetiedot. bulgaricus määritettiin fenotyyppianalyysillä, Pulssikenttäelektroforeesigenotyypityksellä (Pfge) ja PCR-menetelmillä. 45°C: n lämpötilassa kasvaneet viljelmänäytteet olivat sauvanmuotoisia, tuottivat D-laktaattia, tuottivat 1065 emäsparin DNA-fragmentin alukkeiden LB1/LLB1 kanssa ja osoittivat proteolyyttistä aktiivisuutta. 665 kasvinäytteestä L. delbrueckii subsp. bulgaricus ja tai S. thermophilus eristettiin, joista suurin osa tuli Bulgarian Blagoevgradista. (12)
1) F., Felis, Giovanna E., Castioni, A., Torriani, S., and Germond, J. ”Lactobacillus delbrueckii subsp. indicus subsp. Marraskuuta., eristetty intialaisista maitotuotteista”. 2005. International Journal of Systematic and Evolutionary Microbiology. Osa 55. s. 401-404.
2)
3) J., Lapierre, L., Delley, M., Mollet, B., Felis, G., and dellaglio, F. ” Evolution of the Bacterial Speies Lactobacillus delbrueckii: A Partial Genomic Study with Reflections on Prokaryotic Species Concept”. Molekyylibiologia ja evoluutio. 2003. Osa 20. s. 93-104.
4)
5) M., Callegari, M., Ferrari, S., Bessi, E., Cattibelli, D., Soldi, S., Morelli, L., Feuillerat, N. ja Antoine, J. ”Survival of Yogurt bacteries In The Human Gut”. Soveltava Ympäristömikrobiologia. 2006. Osa 72. s.5113-5117.
6)
7) S., Drescher, K., and Heller, K. ”Survival of Lactobacillus delbrueckii subsp. bulgaricus ja Streptococcus thermophilus Fistulated Gottingen Minipigien terminaalisessa ileumissa. Soveltava ja Ympäristömikrobiologia. 2001. Osa 67. s.4137-4143.
8) C., D. Atlan, B. Blanc, R. Portailer, J. E. Germond, L. Lapierre ja B. Mollet. 1996. ”A new cell surface proteinase: sequencing and analysis of the Prtb geeni from Lactobacillus delbruekii subsp. bulgaricus (oik. Journal of Bacteriology. 1996. Osa 178. 3059-3065.
9)
10) Center for Biotechnology Information (NCBI) Genome. Lactobacillus delbrueckii subsp. 11842, täydellinen genomi.
11) de Guchte, M., Penaud, S., Grimaldi, C., Barbe, V., Bryson, K. ja muut. Lactobacillus bulgaricuksen täydellinen genomisekvenssi paljastaa laajan ja jatkuvan reduktiivisen evoluution. PNAS. 2006. Osa 103. s.9274-9279
12)M., Minkova, S., Kimura, K., Sasaki, T., and Isawa, K. ”Isolation and characterization of Lactobacillus delbrueckii ssp. bulgaricus and Streptococcus thermophilus from plants in Bulgaria”. FEMS Microbiology Letters. 2007. Osa 269. s. 160-169.
13)
Edited by Maryruth Penetrante student of Rachel Larsen
Edited by KLB